としている。
また、サイト「Brazil Covid variant could cause reinfection in 61 per cent of people」では、
①これまでのブラジル循環株よりも1.4〜2.2倍感染性が高い。
②従来株に感染した人の防御抗体の 25~61%を突破し再感染
③現行のワクチンのP.1変異株に対する有効性はある程度は保たれている。
としている。

ジャクソンホールにおける植田日銀総裁の講演の抄訳が、日本銀行のサイトから発表されました。
リンクはこれです。
https://www.boj.or.jp/about/press/koen_2023/data/ko230828a1.pdf
抄訳でも結構長いので、当方で、1200文字程度に要約しました。
下記です。
パネルセッション「変曲点にあるグローバリゼーション」 植田日銀総裁
1.アジアの貿易と直接投資の変動
日本からの視点と地政学的緊張の影響についての考察。
1970〜80年代の日米貿易摩擦の例やIMFの分析を引用しての展望。
私(植田日銀総裁)は貿易論の専門家ではないが、日本からの視点を提供したい。アジアの貿易や直接投資の状況が、地政学的緊張の影響を受けて変化していること、特に日本は生産拠点を中国から他のアジアや北米、日本へとシフトしている動きが見られる。この変化はグローバリゼーションの影響の一部として、また地政学的リスク対策として見ることができる。1970〜80年代の日米貿易摩擦を例に、貿易関係の変動が経済の構造変化をもたらすこと、例えば、日本企業が米国での生産を始めたことなどが挙げられる。「地政学的な問題を受けた世界経済の分断化リスク」について、IMFは米国圏と中国圏の間での直接投資規制が発動すると、世界GDPが大幅に減少し、特に東南アジアが深刻な影響を受けると分析している。
2.日本の貿易・投資の動向
日本の輸出動向、対外直接投資のトレンド、日本政策投資銀行のアンケート結果を基にした分析。
日本企業の生産拠点のシフトや国内生産能力増強に関する考察。
日本の輸出動向を見ると、中国向け輸出シェアはコロナ後に一時増加したが、米国やその他アジア向けは安定しており、ベトナムやインドへの輸出は増加している。
日本の対外直接投資では、中国向けは停滞している一方、北米やベトナム、インド向けは増加している。
日本政策投資銀行のアンケートによれば、日本企業が重視する投資先として北米、中国、ベトナム、タイ、インド、インドネシアが挙げられる。
日本企業は、中国からASEANやインド、北米への生産拠点の多様化を進めており、これは地政学的考慮だけでなく、現地の需要増や米国の産業政策の影響も背景にある。
日本企業の国内生産能力増強の動きが目立つが、これは海外生産能力の犠牲によるものではない。
7月の輸出データでは欧州や米国向けが強く、アジア向けは一部減少。
特にハイテク財輸出で中国向けは減少しており、理由についてはわからない。
3.アジアの経済動向とグローバルな影響
アジアの生産拠点の動きや地域経済統合、中国の戦略に関する検討。
中国と米ドルの役割やアジア新興国の地政学的変動に関する展望。
複数の業種でアジアにおける生産能力増強が予定されているが、半導体関連では国内強化が顕著である。
アジアの生産拠点の動きが変わる「変曲点」が近いかもしれない。
アジア新興国の地域経済統合は進行中で、グローバル化の動きは持続している。
中国はアジアだけでなく、他地域への生産移管を進め、人民元を戦略的に貿易金融に使用しており、米ドルの役割に変動が見られるかもしれない。
4.日本経済の現状と中央銀行の課題
本年の日本経済のGDP成長率や物価動向、日本銀行の金融政策についての考察。
地政学的リスクや脱グローバル化の動きなどの不確実要因に関する検討。
本年の日本経済は、第1四半期のGDP成長率が3.7%増でスタートしている。
この強さは、感染症制約の緩和やインバウンド需要の復活によるものである。
第2四半期も6%成長を記録し、個人消費や設備投資が引き続き好調。
物価面では、輸入物価の上昇が国内物価に波及し、7月には3.1%になった。
しかし、年末に向けては低下が予想される。
日本銀行は、2%の物価安定目標に達していないため、金融緩和を継続する。
一方、製造業の国内生産強化や半導体産業への投資が経済を刺激している。
しかし、中国経済の減速や地政学的リスク、脱グローバル化の動きなど、多くの不確実要因が経済見通しを複雑化させている。
中央銀行はこれらの要因を考慮しながら、適切な政策決定の難しさに直面している。
感染症関連の供給ショックも継続して影響を及ぼし、中央銀行は過去の経験を踏まえ、適切な対応を模索中である。
以上
#変曲点

昨晩(2023/08/25)、ジャクソンホールでのパウエルFRB議長の講演要旨を五分の一にまとめてあります。
なお、パウエルさん講演全文テキストは、下記URLサイトでみることができます。
https://www.federalreserve.gov/newsevents/speech/powell20230825a.htm
また、当日の講演ビデオは、下記RLで見ることができます。
https://www.youtube.com/KansasCityFed
以下はパウエル講演要旨です。
ジェローム・H・パウエル議長はジャクソンホールでの経済政策シンポジウムで講演し、インフレ率を目標の2%に引き下げるというFRBのコミットメントを改めて強調した。
FRBはすでに過去1年間政策を引き締めており、自信を持ったインフレが一貫して目標に向かって進むまでさらに利上げする用意がある。
当初は高い需要とパンデミックによる供給の制約によりインフレが急上昇した。
2022年3月までに、インフレを抑制するにはパンデミック関連の需給混乱の緩和と金融引き締めの両方が不可欠であることが明らかとなった。
米国の総合インフレ率は2022年6月に7%でピークに達し、7月までに3.3%に低下した。
変動の激しい食品とエネルギー価格を除いたコアPCEインフレ率は2022年2月に5.4%でピークに達し、7月までに4.3%に低下した。
需要の変化や半導体不足などの供給制約の影響を受ける自動車セクターなどの要因は、インフレが直面する課題を浮き彫りにしている。
全体として、インフレ抑制の一貫した進展を確実にするためには、さらなる金融制限措置が必要であると考えられる。
住宅セクターでは、金融政策の効果がリフトオフ後に明らかになり、2022年には住宅ローン金利が倍増し、住宅着工数、販売数、価格の上昇が減少した。
住宅サービスのインフレがこうした変化を示すまでには時間がかかったが、現在は市場家賃の下落の影響を受けて低下している。
コアPCE指数の半分以上を占める非住宅サービスは、リフトオフ後に大きなインフレ変化を示していない。
このセクターの将来の安定は、価格の安定にとって極めて重要だ。
将来的には、抑制的な金融政策がインフレ管理において大きな役割を果たすことになるだろう。
2%のインフレ率を達成するには、トレンドを下回る経済成長と労働市場の軟化が予想される。
金融環境が引き締まり、通常は経済活動の成長が鈍化しているにもかかわらず、GDP成長率は堅調で、個人消費も引き続き堅調だ。
労働市場は依然としてリバランスを続けており、労働力の供給と需要の両方が変化している。
賃金圧力は緩和し、名目成長率が鈍化しているにもかかわらず、実質賃金の伸びは増加している。
中心目標は引き続き2%のインフレ目標である。
これを達成することは困難であり、特に必要な金融政策の正確なレベルを決定することは困難だ。
金融引き締めが経済とインフレに与える正確な影響と期間は依然として不確実である。
現在の経済状況には、特にインフレと労働市場の動向に関して明らかな不確実性がある。
たとえば、求人数は減少しているものの、失業率は上昇しておらず、労働需要が異常に高いことを示している。
さらに、インフレは現在、以前よりも労働市場の状況により影響を受けているようだ。
こうした不確実性により、金融引き締めの適切な強度を決定することが困難になっている。
少なすぎると目標を上回るインフレが持続する可能性があり、多すぎると経済に悪影響を与える可能性がある。
これへの対応としては、政策調整を決定する前にさまざまなデータを評価する慎重なアプローチが取られるだろう。
最終的な目標は物価の安定であり、これはすべての人に利益をもたらす強力な労働市場にとって極めて重要だ。
以上

[embedyt] https://www.youtube.com/watch?v=Aox8NFGzBNU[/embedyt]
Prime Minister Kishida announced plans to commence the release of treated water from the Fukushima Daiichi Nuclear Power Plant on August 24, 2023. This decision stems from concerns linked to the plant’s shutdown following the 2011 East Japan Earthquake. The ALPS facility, designed to treat radioactive-contaminated water, has been accumulating water in tanks, leading to potential delays in the plant’s decommissioning schedule.
In a 2016 report by the Tritium Water Task Force, five treatment methods were proposed. Due to feasibility, the options of releasing water vapor and discharging into the ocean were chosen. Initial estimates suggested that releasing into the ocean would be the most economical. However, recent estimates from Tokyo Electric Power Company (TEPCO) indicate that the cost for ocean discharge has risen from an initial 3.4 billion yen to 59 billion yen. This is mainly attributed to unexpected costs associated with constructing an undersea tunnel for the discharge and increased expenses to combat potential reputational damage.
On the topic of reputational damage, both TEPCO and Prime Minister Kishida emphasize the need for measures post the oceanic release of the ALPS-treated water. However, the term “reputational damage” has faced criticism. Detractors argue that the phrase implies that there’s harm due to unwarranted negative perceptions, suggesting that the government and TEPCO might be adopting a victim stance.
Assessment of risks associated with reputational damage remains ambiguous. There are concerns that the use of the term by the government and TEPCO might underestimate the actual risks. Particularly, potential broader ecological impacts and cascading risks from oceanic discharge might not be fully accounted for. On the other hand, releasing as water vapor is seen as a more localized method of managing contaminants, with a more limited risk scope.
In summary, the water release plan from the Fukushima Daiichi Nuclear Power Plant is surrounded by extensive debates and concerns. Significant discrepancies between initial and current cost estimates, especially the surge in costs to address reputational damage, could impact the direction and approach of future plans.
はじめに
2020年秋口までの新型コロナウイルスの変異の過程については、昨年11月9日に書いたブログ「近時の新型コロナウイルスの変異株への懸念~「20A.EU1」株と「クラスター5」株の出現~」に記載した。
この時点に記載した主な変異は「D614G」変異が中心だったが、これまでの「D614G」変異を引き継ぐ形で、B.1.1.7系統変異があらわれ、その後も、昨年秋以降、そのほかの新型コロナウイルス変異株は、日々、世界で発見されている。
1.最近2020年秋以降に発見された、いくつかの新型コロナウイルス変異株
昨年11月以降に発見された新型コロナウイルスの変異株には次のものがある。
①イギリス由来変異株
「B.1.1.7」系統の「VUI-202012/01」(下記地図のマゼンタ色地で発見)
②南アフリカ由来変異株
「B.1.351」系統の「501.V2」(下記地図の黄色地で発見)
③ブラジル由来変異株
③-1 「P.2」系統の「20B/S.484K」または「B.1.1.248」(下記地図の濃緑地で発見)
③-2 「P.1」系統のブラジル&日本由来変異株「20J/501Y.V3」(下記地図の薄緑色地で発見)
④アメリカ由来変異株
④-1.オハイオ由来「20G/501Y」(下記地図の空色地で発見)
④-2.テキサス由来「20C-US」(下記地図の空色地で発見)
④-3.カリフォルニア由来「Cal.20C」
⑤ナイジェリア由来変異株「B.1.1.207」(下記地図の橙色地で発見)
2.変異株はいつどこで発見されたのか?変異箇所はどこか?ウイルス特性は何か?
(1)どこの変異か?
一口に変異といっても、それが、新型コロナウイルスのスパイク蛋白質での変異なのか?そうでない部分での変異なのか?によって、ワクチンの有効性という観点からは、意味が異なってくる。
なぜなら、現在開発の新型コロナウイルスワクチンのほとんどが、ウイルスのスパイク部分をターゲットにしたものであるからである。
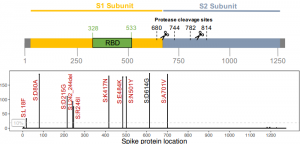
スパイクタンパク質の受容体結合ドメイン(RBD)は配列順位1から1273まである。
スパイク蛋白質はS1とS2の、二つのサブユニットからなる膜貫通タンパク質てあり、N末端のS1サブユニット部分とC末端のS2サブユニット部分とからなる。
S1 (1–685)には細胞表面受容体認識を担う部分があり
S2(686–1273)には膜融合に必要な要素がある。
このS1とS2との間を切断するのが、別のプロテアーゼのフューリン(Furin)である。
この切断部分を、開裂部位(685と686との間)という。
その後S1が新型コロナウイルスの受容体であるACE2受容体に結合する。
S2は、ヒト細胞表面の呼吸器上皮などに発現しているタンパク分解酵素によって切断され、その結果として膜融合が進行する。
なお、たとえば「D614G変異」という場合、その意味するところは
「スパイクタンパク質のアミノ酸残基配列(1から1273)のうちの614番目のアスパラギン酸(D)がグリシン(G)に置換わる変異」
ということである。
(2)これまで確認されている変異株の概要
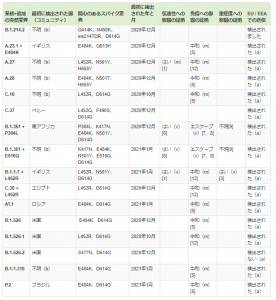
A,イギリス由来変異株「B.1.1.7、20I 」または「VOC202012 / 01」または「VUI-202012/01」または「 501Y.V1」
(A).いつ、どこで発見されたのか?
最初、2020年9月20日にイギリス南東部ケントで症例発見。10月初旬にゲノム配列公開。
グレーターロンドンにも9月21日に出現。
2020年12月20日以降、米国やカナダなどでも、この変異株による症例発生。
以来世界各地で感染報告があり、現在62か国で発見
英国外の感染地としては
ポルトガル、イスラエル、ベルギー、オランダ、オーストラリア、デンマーク、フランス、アメリカなど。
D614Gよりもはるかに感染性が高いと考えられているB.1.1.7株は、
(B).変異箇所は?
The K variants of Covid-19 carry 17 non-silent mutations and 8 in the Spike protein.
23の変異をもつ。
ウイルスゲノムに17の変異があり、うち、8がS蛋白質の変異である。
重要な変異は下記である。
N501Y変異ー感染力の増大化とウイルス量の増加に寄与している。なお、この変異は、南アフリカ変異株でも見られている。
HV 69/70欠失ースパイクタンパク質のコンフォメーション変化(タンパク質の立体配座の形が変わる)につながる可能性がある。
P681HーS1 / S2フューリン切断部位(685と686との間)の近くでの変異である。
スパイクタンパク質の外側のORF8Q27stop変異
ORF8終止コドン(Q27stop)ーORF8の変異であるが、変異による機能変化は不明である。
(C).変異によるウイルス特性の変化は?
N501Y変異はS蛋白質の受容体結合ドメインに位置しており、スパイクRBDとACE2受容体との相互作用により感染力増強(感染速度が速くなる可能性)とウイルス量の増加をもたらしているとされる。
従来ウイルスより50〜70%最大70%速く(感染性が約1.5倍高いともいえる)拡散する。(他変異株より56%伝染性が高い。との数字も出ている。)40%以上増加
3つの欠失(位置は69/70)は、S蛋白質のコンフォメーション変化(タンパク質の立体配座の形が変わる)に寄与している。
実効再生産数R値が+0.4上昇する可能性(R値が0.4から0.7の間)がある。
免疫回避の可能性についての検証は不十分だが、グラスゴー大学教授によると、これがあると、ワクチンの有効性に懸念が生じることにもなるとしている。
(D).その他
今後数か月(2021年3月までに)で世界中で「優勢」になるとの見方もある。
イギリス政府の科学諮問グループNervtag報告(SAGE meeting paper)によると、「B.1.1.7」は、データは限られているが、死亡リスクが1.3倍に増加とのことだが、詳しい検証はまだ得られていない。
英国ジョンソン首相は、この報告を受け、「英国新型コロナウイルス変異体「B117」が従来株より致命的な証拠がある」と語ったことで、米CDCが、その発言の裏づけの資料を求めている。
米CDCのモデリングでは2021年3月迄に英変異株が主流になるとの見通しを立てている。
一方、英国内の専門家には「そう断じるのは時期尚早、ジョンソン首相の発表に驚いた」とする向きもある。
英ウォーリック大学マイク・ティルデスリー博士は、患者1,000人あたり死者数が10人から13人に増えたが「少量のデータに基づくもの」と警告している。
また、WHOのKerkhove博士は、英国変異株の子供へ感染について、感染した子供には無症候性が多く、症状は、子供ほど軽度になる傾向があるので、見逃しによる子供への感染者増には警戒すべき、としている。
「B.1.1.7」新型コロナウイルス変異株が脅威でないと思われる理由として、感染力増強は毒性の低下につながりうるとの「毒性の低下の法則」(セオボールドスミス)を持ち出すのもいるが、
これには異論があり狂犬病とHIVの場合は、宿主を殺しても広がりうると反論する向きもある。
なお、これまでに、この変異株が、重症度やワクチンの有効性に影響を与えることを示唆する証拠はない。
参考
「Emergence of SARS-CoV-2 B.1.1.7 Lineage — United States, December 29, 2020–January 12, 2021」
B.南アフリカ由来変異株「B.1.351」または「501.V2」または「20C / 501Y.V2」
•病原性の増加の証拠はありません
•invitro研究は、自然感染後の免疫回避の可能性とワクチン誘発抗体の効力へのわずかな影響を示唆しています
(A).いつ、どこで発見されたのか?
2020年10月8日に、南アフリカの東ケープ州ネルソンマンデラベイで最初に検出された。
2020年12月下旬にザンビアでも確認、その後、以下記載の各国へ、急速にひろまった。
2021年1月25日現在で、感染が報告された国は、26カ国ある。
南アフリカ、英国、ドイツ、オーストラリア、フランス、ベルギー、ボツワナ、アラブ首長国連邦、スイス、アイルランド、ケニア、フィンランド、韓国、スエーデン、ノルウェイ、ポルトガル、ニュージーランド、オランダ、デンマーク、スペイン
などである。
俗称「Covid VSOP」とも呼ばれている。
(B).変異箇所は?
スパイク(S)タンパク質に21個の変異がある。
主な変異箇所は
S蛋白質(K417N 、E484K、N501Y)
P.1系統orf1b欠失del11288-11296
スパイクたんぱく質受容体結合ドメインの3つ(K417N、E484K、N501Y)の変異が特徴である。ORF1b欠失
(C).変異によるウイルス特性の変化は?
重要な3つの変異(N501Y変異、E484K変異、K417N変異)によるウイルス特性変化(感染の増加とウイルス量の増加に関連か?)に懸念がもたれている。
すなわち
N501Y変異ー感染性が高い原因、体内ウイルス量を多くする。これは英国変異株にもある変異。
E484K変異ー免疫応答回避の特性をもち、中和抗体の生成を10分の1低下させる可能性。
K417N変異ーE484K変異同様、現在のワクチンでの誘導抗体が亜種に結合し中和しない懸念がある。
これら二つの変異(E484K変異とK417N変異)はいずれもスパイクタンパク質での変異である。
一方、現在開発接種中の主力ワクチンはいずれもスパイクタンパク質を標的としており、これら2つの変異によって、主力ワクチンの有効性が左右されるのではないか?ワクチン誘導抗体が変異種に結合し中和しない可能性があるのではないか? との懸念を、専門家は持っている。
特に、E484K変異が、一部のポリクローナル抗体およびモノクローナル抗体による中和に影響を与える可能性があることを示すいくつかの証拠が出ている。
なお、この変異株には、英国変異株「B.1.1.7」が有する変異である「69/70欠失」は含まれていない。
現在、この変異が病気の重症度に影響を与えることを示唆する証拠はない。
(D).その他
南アフリカで発見の新型コロナウイルスの変異株「501Y.V2」について専門家は下記の懸念を持っている。
①若者に対する感染しやすさがあるのでは?
②再感染の懸念があるのでは?
③ヒト→ヒト感染が容易なのではないか?
④開発ワクチンに対する耐性が、やや高いのではないか?
これについて、昨年末南アフリカ新型コロナウイルス変異株「501Y.V2」特定のNGS-SA等科学者達は501Y.V2特性究明のためのコンソーシアムを結成し、当該変異株に対する当面の課題として以下の点を上げた。
①ワクチン接種者の血液で、ワクチン誘導抗体の効果を検証
②501Y.V2に対してワクチンが機能しない可能性を追及
なお、新型コロナウイルスの南アフリカ変異株(501Y.V2)の免疫応答回避メカニズムについては、下記のネイチャーのニュース
「Fast-spreading COVID variant can elude immune responses」
がある。
C.ブラジル由来変異株「20B/S.484K」または「B.1.1.248」
(A).いつ、どこで発見されたのか?
7月に最初に出現し、10月に最初に検出。
12月中旬にブラジル北部のアマゾナス州のマナウスから検出。
「Brazil(ian) variant」ともいわれる。
(B).変異箇所は?
27の変異があり、そのうち11はスパイク蛋白質の変異であ利、スパイク蛋白質外に16の変異がある。
主要なスパイクたんぱく質の変異としては
N501Y変異
E484K変異
がある。
(C).変異によるウイルス特性の変化は?
(D).その他
E.ブラジル&日本由来変異株「20J/501Y.V3」また「P.1系統」
1月の日本の空港でのスクリーニング中にブラジルからの旅行者で発見されました。現在、ブラジルのアマゾナス州で広く流通していることが知られており、フェロー諸島、韓国、米国でも観察されています。
•伝染性および/または病原性への影響は不明
•再感染の逸話が報告されていますが、免疫回避の可能性は未解決のままです
(A).いつ、どこで発見されたのか?
B.1.1.28系統からの分岐株と見られる。最近検出された3つのSARS-CoV-2系統(B.1.1.7、B.1.351、およびP.1)
P.1は、B.1.1.28系統の子孫ともいえる。
この亜種の最初の系統は、東京郊外の羽田空港での定期的なスクリーニング中に検査されたブラジルからの4人の
テストしサンプルの42%にウイルス検出。
マナウスに旅行した人々からも検出。
N501YおよびE484K変異を持つP.1として識別された新しいバリアントは、12月15〜23日にマナウスで収集された31の陽性PCRテストサンプルのうち13で検出されました。3月から11月の間に見られたゲノム監視サンプルには存在せず、それがどれほど迅速に出現して再現されたかを強調しています。
P.1コロナウイルス変異体(株をより伝染性にし、COVID生存者をよりよく再感染させ、ワクチンの効果を低下させる可能性のある少なくとも2つの重要な変異を持っている)
P.1バリアントはこれまでブラジルでのみ確認されており、ブラジルからの旅行者(主にアマゾナス州から)で日本と韓国で報告されています。アマゾナス州の州都マナウスでは、現在COVID-19の症例が急増しており、医療システムに大きな圧力をかけています。
日本の厚生労働省は、2021年1月2日にブラジルのアマゾナス州から来日した4人の旅行者から、新しい変異体検出と、2021年1月10日に発表。
日本の国立感染症研究所(NIID)は、新しい変異体には12の変異があり、そのうちの1つ(「N501Y」変異のことか?)は英国と南アフリカで見つかった変異体にも存在すると、ブラジルの保健省に伝達した。
1月25日、アメリカのミネソタ州で始めてP1変異株を確認した。
(B).変異箇所は?
スパイクタンパク質のN501Y、E484K、およびを含む17個のアミノ酸の変化。
B.1.1.248系統と同じく、スパイクタンパク質にN501YとE484KとK417Nを含む17の変異がある。
スパイク蛋白質での変異としては、
:L18F, T20N, P26S, D138Y, R190S, K417T, E484K, N501Y, H655Y, T1027I
がある。
スパイクタンパク質の外側の ORF1b欠失
ORF1ab:F681L, I760T, S1188L, K1795Q, del3675–3677 SGF, E5662D0352
ORF3a: C174G
ORF8: E92K
ORF9: Q77E
ORF14: V49L N: P80R
(C).変異によるウイルス特性の変化は?
(D).その他
系統B.1.1.248
ウィキペディアから、無料の百科事典
ナビゲーションにジャンプジャンプして検索
口語的にブラジル(ian)バリアントとして知られている系統B.1.1.248 [1] [2] [3]は、COVID-19を引き起こすウイルスであるSARS-CoV-2のバリアントの1つです。[4] SARS-CoV-2の[5] [6]このSARS-CoV-2の亜種は、2021年1月6日に国立感染症研究所(NIID)によって、4日前にブラジルのアマゾナスを訪れて東京に到着した4人で最初に検出されました。[3] [7 ] 2021年1月12日、ブラジル-英国CADDEセンターは、アマゾン熱帯雨林の最大の都市であるアマゾナス州マナウスで、P.1の新しい系統の13の局所症例を確認しました。[5]新しい系統は、マナウスからの3月から11月のサンプルには存在しませんでしたが、同じ都市で収集された2020年12月のサンプルの42%で識別され、最近の頻度の増加を示唆しています。[5]
Carolina MVoloch他による論文のプレプリント。 2020年にシーケンスされたブラジルのサンプル間の関係について説明し、ブラジルで流通しているSARS-CoV-2の新しい系統「B.1.1.248」を特定しました。これはB.1.1.28に由来します。 7月に最初に出現し、10月に最初に検出されたと説明されていますが、公開(2020年12月)の時点では、頻度は大幅に増加していますが、それでも大部分(ケースの61%)は州都リオデジャネイロに限定されていました。 。[8] 5月には、サンプルの大部分が系統B.1.1.33でしたが、9月までにB.1.1.28が大幅に広がり、10月と11月には、それぞれ新しい変異系統(B.1.1.248)が優勢でした3。パンゴリンツールを使用した他の4つの分類。[9]このホワイトペーパーでは、変更E484K(B.1.1.28とB.1.1.248の両方に存在)がサンプル全体に「広く広がっている」(たとえば、1セットの38サンプルのうち36サンプル)と特定しています。[9]
.1系統(別名20J / 501Y.V3)
P.1バリアント(B.1.1.248としても知られています)は、いくつかの理由で科学者に関係しています。それは、それをより危険にする可能性のある2つの注目すべき突然変異があることから始まります。
第一に、それは変異体をより感染性にするかもしれないスパイクタンパク質突然変異を持っています。N501Y(または「ネリー」)として知られるこの変異は、英国を急速に追い抜いているB.1.1.7バリアント、およびB.1.351(501.V2またはN501Yとしても知られる)にも存在します。 V2)南アフリカで出現したバリアント。この変異により、ウイルスのスパイクタンパク質がヒト細胞とより容易に結合できるようになり、感染性が高まる可能性があります。研究によると、B.1.1.7バリアントは、以前は優勢だった菌株よりも50%以上感染しやすい可能性があります。そのため、科学者や公衆衛生当局はそれについて非常に心配しています。
P.1バリアントには、E484Kとして知られる「エスケープ変異」もあります。これは、南アフリカのB.1.351バリアントにも存在し、実験室での実験で、コロナウイルスが以前の感染によって生成された防御抗体を回避するのに役立つことがわかっています。抗体薬の影響を受けにくい。言い換えれば、コロナウイルスの以前の株にすでに感染している誰かがこの突然変異を持つ変異体によって再感染する可能性があり、突然変異はコロナウイルスがCOVIDワクチンによって生成された抗体を回避することを可能にするかもしれません。
明確にするために、これは、現在利用可能なコロナウイルスワクチンがE485K変異を持つ変異体に対して機能しないことを意味するものではありません。現在米国での使用が許可されている2つのワクチンのメーカーであるModernaとPfizer-BioNTechは、ラボテストで、南アフリカのB.1.135バリアントに対してmRNAワクチンの効果がわずかに低いことを発見し、両社はすでに取り組んでいます。問題に対処するためにワクチンを更新します。他の場所では、多くのコロナウイルスの専門家は、ワクチンがE484K変異を持つこれらの新しい株に対して同等の防御を提供することを期待しています。より多くの研究が必要ですが、今のところ、それは最悪の場合の現実世界のシナリオのようです ワクチンは変異体による重篤な病気を予防しますが、軽度または無症候性の感染症の予防には効果が低い可能性があります。
また、ワクチンは、この変異や他の変異を考慮して更新することができます。
P.1とB.1.351バリアントも別の変異K417を共有していますが、それがウイルスにどのように役立つかについてはあまり理解されていません。
関心のある変異体が地理的に異なる別々の場所で同時に
In Brazil, it’s too early to conclude whether a variant now circulating there, called P.1, is inherently more transmissible. First reported on January 12 in the state of Amazonas, it’s been associated with a devastating surge in cases in Manaus, a city where researchers had previously estimated that 75 percent of residents had already been infected with SARS-CoV-2. But it’s still unclear whether properties of the virus itself are contributing to the surge, says virologist Paola Resende of the Oswaldo Cruz Institute in Rio de Janeiro. “In Brazil, we can see a lot of parties, we can see the pubs crowded, and people are on the streets not wearing masks. I think this behavior of the population is the main reason [for] the increase.”
There’s less information on the P.1 variant, which health officials in Minnesota reported January 25 has been detected there, marking the first observation in the US. Because its mutation pattern is similar to B.1.351—namely, it shares the E484K and K417N RBD mutations—“there would be reasons to believe that what applies for one would likely apply to the other,” Moore notes.
Resende and her colleagues have recently documented two cases when people became reinfected with a new variant. In one, the reinfection was caused by P.1. The other incident of reinfection was caused by P.2, a closely watched emergent sister variant that carries fewer changes overall but harbors the N501Y and E484K mutations. Given that reinfections are known to occur with SARS-CoV-2, albeit rarely, such anecdotal observations don’t tell researchers if it’s more likely to happen with the new variants. Nevertheless, “all mutations located in the receptor binding domain, we need to pay attention to,” Resende says.
Researchers are still investigating the effect of the P.1 variant’s vulnerability to vaccines in Brazil, Resende says. As for the B.1.351 variant, a second experiment in Hatziioannou’s study provides some insights. She and her colleagues examined antibody-containing plasma from 20 people who had either received the Moderna or the Pfizer/BioNTech vaccine. The team tested the plasma against the dominant SARS-CoV-2 spike protein and pseudoviruses engineered to have the variant’s RBD mutations, either individually or in combination. The antibodies proved significantly less effective in neutralizing the pseudoviruses compared to pseudoviruses with the original spike protein, with a one- to threefold decrease in antibody potency. “It’s a really small difference,” she says, adding that it’s not entirely clear why the South African team—testing antibodies from survivors of natural infections against the actual RBD protein—observed a more dramatic drop in antibody potency.
|
P.1, 20J/501Y.V3 |
Discovered in travelers from Brazil during screening at a Japanese airport in January; now known to widely circulate in Brazil’s Amazonas state and also observed in the Faroe Islands, South Korea, and the US |
17 amino acid changes, including N501Y, E484K, and K417N on the spike protein; ORF1b deletion outside the spike protein |
• Effect on transmissibility and/or virulence unknown • Anecdotes of reinfections reported, but potential for immune evasion remains unresolved |
P.1には約20の変異があり、そのうち17の固有のアミノ酸の変化、3つの欠失、および挿入があります。これらのうちの2つは、以前の話で詳しく説明しましたが、N501YとE484Kです。これは、受容体結合ドメインで発生し、伝達率の増加に関連するスパイク変異です。ORF1bの欠失に加えて、P.1には、いわゆるUKバリアントであるB.1.1.7、およびいわゆる南アフリカバリアントである501.V2と共通するこれらの変異があります。3つすべてにおいて、これらの突然変異は独立して発生しました。これは、単独で、または他の突然変異と組み合わせて、ウイルスに何らかの利点をもたらし、生存の可能性を高めることを示している可能性があります。
P1は、アマゾニア州滞在後、日本に帰国した人から発見され、日本から保健省に通達があった。その後、12月に採取したマナウス市在住の患者のサンプルから同型変異種が検出された。P1には、英国や南アフリカで見つかった感染力がより強い変異種と同じ変異が見られ、従来型ウイルスより感染を引き起こし易いとみられている。
最初の感染からの中和抗体がP.1変異体(ブラジル)を認識しない可能性があるという初期の証拠もいくつかあり、これは再感染、さらにはワクチンの失敗につながる可能性があります。私たちはこれを注意深く見守らなければなりません」とラムは言いました。
F.アメリカ由来変異株
F-1.オハイオ由来「20G/501Y」
COH.20G / 501Y
N501Y変異の2例は、オハイオ州コロンバスで12月下旬以降、および他の米国の州で検出されています。
ウイルスのスパイクタンパク質にあるN501Y。英国で特定されたB.1.1.7バリアントに存在する他のほとんどの変異を欠いています。
•伝達性、病原性、および/または免疫回避の変化の証拠はまだありません
A.いつ、どこで発見されたのか?
B.変異箇所は?
C.変異によるウイルス特性の変化は?
D.その他
F-2.テキサス由来「20C-US」
A.いつ、どこで発見されたのか?
B.変異箇所は?
C.変異によるウイルス特性の変化は?
D.その他
F-3.カリフォルニア由来「Cal.20C」
A.いつ、どこで発見されたのか?
B.変異箇所は?
C.変異によるウイルス特性の変化は?
D.その他
S Q677H、
「中西部」バリアントと呼ばれることもあります
S Q677H変異を含むウイルスは、最近、オハイオ州で12月と1月に分析されたサンプルで頻繁に発生し、中西部の複数の州でも発見されています。
スパイクタンパク質のQ677H変異、Mタンパク質のA85S、およびヌクレオカプシドタンパク質のD377Y
•これまでのところ、伝達性、病原性、および/または免疫回避の変化の証拠はありません
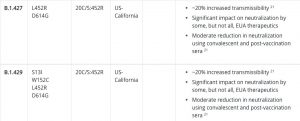
L452R、B1429
L452R変異自体は、昨年、米国とヨーロッパで観察されました。2021年1月、カリフォルニアの複数の郡で頻度が急速に上昇しました。
スパイクタンパク質にあるL452R変異
カリフォルニアでのいくつかの大発生に関連していますが、サージがバリアント自体によって引き起こされているかどうかは不明です。
•調査中のワクチンの有効性への影響
参考
「Emerging SARS-CoV-2 Variants」
3.憂慮すべき免疫応答回避メカニズムをウイルスにもたらす変異
①英国の変異種(B.1.1.7)
②南アフリカ由来変異株
「B.1.351」または「501.V2」(下記地図の黄色地で発見)
12月末現在英国、フィンランド、スイス、日本、オーストラリアの5か国で発見
①
スパイク遺伝子のみ標的としている現在の開発ワクチンでは限界。
多様な免疫応答にターゲットも
発見
南アフリカ、英国、ボツワナ、ザンビア、豪、独、アイルランド、仏、スイス、韓国、スウェーデン、ノルウェー、フィンランド、オランダ、日本等
③ナイジェリア由来変異株
「P681H」(下記地図の橙色地で発見)
三番目に発見の変異株
ナイジェリアでも、変異株の報告が。
④ブラジル&日本由来変異株
「20J/501Y.V3」(下記地図の薄緑色地で発見)
四番目に発見の変異株
⑤ブラジル由来変異株
「20B/S.484K」「B.1.1.248」(下記地図の濃緑地で発見)
五番目に発見の変異株
「厚生労働省はブラジルから到着した4人から、英国や南アフリカで報告されたものと異なる新たな新型コロナウイルス変異種が検出と発表
変異株はB.1.1.248系統に属し、S蛋白質に変異12箇所。
変異株VOC-202012/01や501Y.V2と同様、S蛋白質受容体結合部位にN501Y変異の外、501Y.V2と同様E484K変異」
日本の国立感染研究所からの日本で発見新型コロナウイルス変異株の報告
https://www.niid.go.jp/niid/en/2019-ncov-e/10108-covid19-33-en.html…
①感染研情報はゲノム配列データに限定
②変異株の感染性、病原性、ワクチンの有効性への影響評価にはさらなる調査が必要。
③変異はB.1.1.248系統。N501YとE484Kを含む12
④ブラジル株とは異なる
⑥アメリカ由来変異株
ア.オハイオ由来
「株名不明」(下記地図の空色地で発見)
六番目に発見の変異株
イ.オハイオ由来
「20G/501Y」(下記地図の空色地で発見)
七番目に発見の変異株
ウ.テキサス由来
「20C-US」(下記地図の空色地で発見)
八番目に発見の変異株
夫々の発生場所は以下の地図ご参照
エ、カリフォルニア由来
変異箇所は「L452R」
最初3月にデンマークで確認、カリフォルニアでは5月に。
L452Rは2020年11月下旬から12月上旬間ではテストサンプルの約3.8%、12月下旬から2021年1月上旬にかけては25.2%以上に増加している。
UCSF(カリフォルニア大学サンフランシスコパルナサスキャンパス)が注目しているのは下記の二点のようである。
●カリフォルニア変異株には、S蛋白質に3つの変異。
●うち1つは、英国のとは異なる変異であることに注目。
新型コロナウイルスはACE2受容体との相互作用により、呼吸上皮細胞に感染の可能性が。
S蛋白質はS1とS2の2サブユニットからなる膜貫通タンパク質。
S1 (1–685)には細胞表面受容体認識を担う部分が。
S2(686–1273)には膜融合に必要な要素が。
L452R変異はS1部分変異。
中和抗体の影響を受けにくく
C.ナイジェリア由来変異株「B.1.207」
(A).いつ、どこで発見されたのか?
変異株は8月3日と9月9日収集サンプルのうち、ナイジェリアのオスン州の2人の患者から発見された。
この変異株は「P681H」変異を有している。
「B.1.1.7」との共通変異があるかどうかは、まだ確認されていない。
(B).変異箇所は?
P681Hスパイクタンパク質の変異をもつ。
(C).変異によるウイルス特性の変化は?
変異箇所(681)が、フューリン切断部位(685と686との間)に直接隣接していることで、ACE2受容体との親和性が増すのではないのか?という点に懸念がもたれている。
(D).その他
なし
参考
1.「Emerging SARS-CoV-2 Variants」
2.「Resurgence of COVID-19 in Manaus, Brazil, despite high seroprevalence」
3.「A Guide to Emerging SARS-CoV-2 Variants」
5.「SARS-CoV-2変異株B.1.1.7由来のスパイク変異体コレクション」
6.「COG-UK report on SARS-CoV-2 Spike mutations of interest in the UK」
目次
はじめに
1.最近2020年秋以降に発見された、いくつかの新型コロナウイルス変異株
2.変異株は、いつどこで発見されたのか?変異箇所はどこか?ウイルス特性は何か?
3.懸念される変異はなにか?
(1)懸念される共通の変異
(2)免疫応答回避メカニズムはあるのか?
(3)変異によるウイルスのコンフォメーション変化(蛋白質の立体構造変化)はあるか?
(4)再感染の可能性はあるのか?
(5)風土病化への懸念、ADEへの懸念はないのか?
(6)変異による死亡率(fatality rates )の上昇の懸念はあるのか?
(7)若年層への感染強化への懸念はないのか?
(8)ワクチン効力の無力化はあるか?
(9)変異株を検出する検査体制の強化はできるのか?
おわりにー大陸を横断しての新型コロナウイルスの同時変異は「収斂進化」の帰結なのか?ー
はじめに
2020年秋口までの新型コロナウイルスの変異の過程については、昨年11月9日に書いたブログ「近時の新型コロナウイルスの変異株への懸念~「20A.EU1」株と「クラスター5」株の出現~」に記載した。
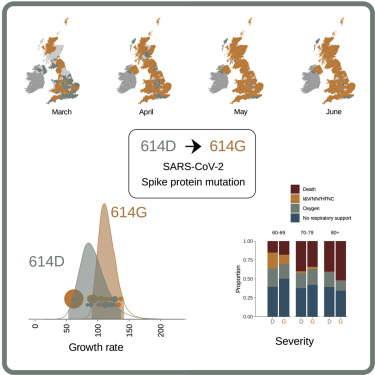
この時点における主要変異は「D614G」変異であった。
D614G変異がすべての始まり
「D614G」変異は、スパイクタンパク質のS1ドメインとS2ドメイン間の接触を妨害し、コンフォメーション(蛋白質の立体構造)の変化(オープン化)により、ウイルスの受容体への結合を強化することで感染力を強化するとされてきた。
S蛋白質はS1とS2の2サブユニットからなる膜貫通タンパク質であり、
S1 (residue1–685)には細胞表面受容体認識を担う部分が。 S2(residue686–1273)には膜融合に必要な要素が
ある。
このS1とS2との間のresidue682からresidue688がフーリン開烈部位である
![]()
「D614G」変異は、ACE2への結合を強化し、人間の気管支における複製を増加させることに寄与した。
D614G変異株は、2020年3月初旬に有意なレベルで最初に検出され、翌月には世界的に優勢な変異株となった。
この武漢株から最初に変異したD614G変異が、その後現れた新型コロナウイルスの多くの変異株に残存して承継されてきたのは、ある意味、脅威でもある。
近時の研究では、2020年1月24日中国でD614G変異が検出されていたとし、米国でのD614G変異の最初の症例は、2020年2月28日にフロリダとロードアイランドで報告されていたとのことである。
その後は、ブログ「近時の新型コロナウイルスの変異株への懸念~「20A.EU1」株と「クラスター5」株の出現~」に記載したように、オランダやデンマークやスペインの農場に、ミンクへの感染の主役として、「クラスター5」と呼ばれる変異株が現れた。
D614G変異からクラスター5変異へ
これは、中和抗体に対する感受性を低下させる懸念が持たれる変異であり、また、ミンクからヒトへの共通感染症が懸念される変異でもあった。
再び、ヒト宿主のウイルスとして新型コロナウイルスが現れたのは、2020年11月初旬のデンマークのミンク農場から採取の214例のヒトコロナウイルスであった。
オランダとデンマークで発生した新型コロナウイルスの塩基配列の多くには、スパイクタンパク質の受容体結合ドメインに「Y453F変異」が見られた。
そして、この「Y453F変異」も、ACE2に対する結合親和性を促す変異であることが確認された。
デンマークで発生した新型コロナウイルスには、3つの変異(①欠失69_70、②I692V変異、③M1229I変異)が確認された。(下記 2-(6)デンマーク由来株「クラスター5」をご参照)
また、新型コロナウイルス感染から回復した患者の血漿からは、ウイルスに対する中和活性の有意な減少が見られた。
しかし、この時点では、この変異が、中和抗体の形成を阻害するとまでは結論付けられなかった。
新しい変異の連鎖への始まり
それまでの「D614G」変異を引き継ぐ形で、「B.1.1.7系統変異」があらわれたのは、9月20日、イギリス南東部のケントであった。
D614G変異は、圧倒的にB.1系統に多かった。
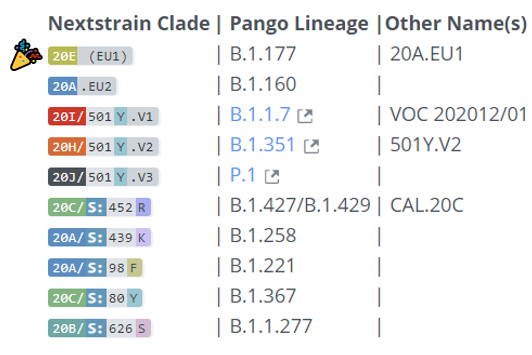
(なお、ここでいう「系統」とは新型コロナウイルスに関して世界共通の系統分類方法である 「Pangolin系統」(Pangolin lineage )のことである。)
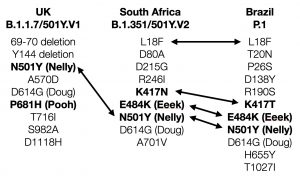
B.1.1.7系統(ケント・バリアント ともいわれる。)は、スパイクタンパク質に、「D614G」を含む、次の変異があった。
「ΔH69/ V70、ΔY144、N501Y、A570D、D614G、P681H、T716I、S982A、およびD1118H」
発見当初、この変異株は「調査中の変異株」(VUI Variant Under Investigation) であるとし、「VUI-202012/01」と命名されたが、感染拡大を見込み、「懸念する変異株」(VOC Variant of Concern)であるとし、「VOC-202012 / 01」または「B.1.1」として再指定された。
それまでの有力変異「D614G」を引き継いだ形で現れた「B.1.1.7系統変異株」は、昨年秋以降、その他の新型コロナウイルス変異株とともに、世界中へと感染を拡大していった。
当時、英国当局は、この変異株には、感染率が70%高くなる可能性があると警告していたが、さらなる研究により、元の武漢ウイルスよりも36〜71パーセント感染性が高い可能性があることが示唆された。
この変異株の出どころはどこなのか?は謎であるが、クイーンズランド大学のウイルス学者KirstyShort氏など、一部研究者の間では、「長い間、新型コロナウイルスと戦い闘病した免疫不全患者が発症の元なのでは?」との推測をしている。
どうして、新しい変異株が、従来の野生株よりも、強い伝染性を持つのか?②ついては謎ではあるが、その一つの原因として「創始家効果」(founder effect)を上げる専門家もいるようだ。
どうして同じ変異が異なる大陸で起きているのか?
これまで、ウイルス変異の理由は2つあるとされてきた。
すなわち
①ウイルス複製過程でのミスコピーによる変異
②ワクチン接種などで抗体産生しウイルスが中和されようとするとき、中和回避のために、ウイルスは変異する。これを免疫エスケープバリアントと呼ぶ。
しかし、これ以外の第三の変異があるのではないかと、専門家は見ている。
すなわち、その後、同時発生的に世界各地で起こった新型コロナウイルスの変異には、共通の変異が見られたことが、専門家の関心を呼んでいるのだ。
つまり、そのことは、新型コロナウイルス側の選択によって、世界バラバラな感染地のもとで、共通の変異が起こっているのではないのか?という懸念でもある。
3つの異なる変異(N501Y変異とE484K変異とK417変異)が、3つの異なる大陸で、異なる系統(Lineage)で出現したにもかかわらず、同じ突然変異をそれぞれ獲得しているのは、偶然ではなく必然の「収斂進化」( convergent evolution)なのではないのか?と専門家は危惧している。
インド・アショカ大学のジャミール氏(Shahid Jameel)は、2021年2月開催のGYANTEEKAのプレゼンテーションで「二つの変異(N501YとE484K)の両方を持つウイルスは、抗体反応回避か、抗体効果低下の可能性があり、また484変異は、アミノ酸の極性変化(Polarization)をもたらす」と話している。
これまでの変異株での主要な変異箇所一覧
1.最近2020年秋以降に発見された、いくつかの新型コロナウイルス変異株
昨年11月以降に発見された新型コロナウイルスの主な変異株には次のものがある。
(1)イギリス由来変異株
(1)-1.「B.1.1.7」系統の「VOC-202012/01」
(1)-2.「B.1.525」系統
(2)南アフリカ由来変異株
(3)ブラジル由来変異株
(3)-1 「P.1」(B.1.1.28.P1)系統の「20J/501Y.V3」(ブラジル&日本由来変異株)
(3)-2 「P.2」(B.1.1.28.P2)系統の「20B/S.484K」
(3)-3 「P.3」(B.1.1.28.3)系統
(3)-4 「VOI N.9」 (B.1.1.33)系統
(4)アメリカ由来変異株
(4)-1.オハイオ由来「COH.20G / 501Y」
(4)-2.テキサス由来「20C-US」
(4)-3.カリフォルニア由来「Cal.20C」または「B.1.429」または「20C/S:452R;/B1429」
(4)-4.中西部由来「Q677H」変異株
(4)-5.中南部&南西部由来「Q677P」変異株
(4)-6.ニューヨーク由来「B.1.526」変異株
(5)ナイジェリア由来変異株
(6)デンマーク由来株
「クラスター5」
(7)インド変異株
(7)-1「N440K」変異(インド南部由来株)
(7)-2.二重変異株「B.1.617」
(7)-3.三重変異株(B.1.617+スパイク蛋白質V382L)
(7)-4.ベンガル変異株(B.1.618)
(8)その他の変異株
(7)-1「Fin-796H」変異(フィンランド由来株)
(7)-2 「B.1.1.316」変異(感染経路不明、日本で発見)
(7)-3.ブラジルでの「B.1.1.33」系統が「E484K」変異をもち再登場
(7)-4.「アンティグア・コビッド」変異株
(7)-5.フランスのブルターニュ地方で発見の変異株「ブルターニュ変異株」(Breton variant)
なお、変異株の特性などについては、下記のサイトを参照
「Coronavirus Variants and Mutations」
「Coronavirus mutations and variants—what do we know?」
2.変異株は、いつどこで発見されたのか?変異箇所はどこか?ウイルス特性は何か?
(1)イギリス由来変異株
イギリス由来変異株の総合的な検証については、
『SARS-CoV-2 variants of concern and variants under investigation in England』(2021年3月11日発表)
をご参照のこと。
(1)-1.「B.1.1.7」系統の「VOC-202012/01」
A.いつ、どこで発見されたのか?
2020年9月20日イギリス南東部ケントで症例発見。このことから「ケント・バリアント」とも呼ばれている。
9月21日にグレーターロンドンでも発見。
2020年12月20日以降、米国やカナダなどでも、この変異株による症例発生。
以来世界各地で感染報告があり、現在62か国で発見。
英国外の感染地としては
デンマーク(2020/12/)、ベルギー(12/)、オランダ(12/)、オーストラリア(12/)、イタリア(12/)、アイスランド(12/)、ジブラルタル(12/)、ウルグアイ(12/20)、シンガポール(12/23)、イスラエル(12/23)。北アイルランド(12/23)、ドイツ(12/24)、スイス(12/24)、アイルランド(12/25)、日本(12/25)、カナダ(12/26)。フランス(12/26)、レバノン(12/26)、スペイン(12/26)、スウェーデン(12/26)、ヨルダン(12/27)。ノルウェイ(12/27)、ポルトガル(12/27)、フィンランド(12/28)、韓国(12/28)、チリ(12/29)、インド(12/29)、パキスタン(12/29)、アラブ首長国連邦(12/29)、米・コロラド(12/29)、マルタ(12/30)、台湾(12/30)。中国(12/31)、ブラジル(12/31)、トルコ(2021/01/01)、ルクセンブルグ(01/02)、ベトナム(01/02)、ギリシャ(01/03)、ジャマイカ(01/03)、キプロス(01/03)、ニュージーランド(01/03)、タイ(01/03)、オーストリア(01/04)、イラン(01/05)、オマーン(01/05)、スロバキア(01/05)、米・カリフォルニア(01/06)、米・フロリダ(01/06)、米・コロラド(01/06)、米・ジョージア(01/06)、米・ニューヨーク(01/06)、ルーマニア(01/08)、ペルー(01/09)、メキシコ(01/10)、ロシア(01/10)、マレーシア(01/11)、ラトビア(01/11)、エクアドル(01/12)。フィリピン(01/13)、ハンガリー(01/13)、ガンビア(01/14)、ドミニカ(01/15)、米・ロサンジェルス(01/16)、アルゼンチン(01/16)、チェコ(01/18)、モロッコ(01/18)、クウエート(01/19)、ナイジェリア(01/25)、セネガル(01/28)、
その他
アルバ、バングラディッシュ、バルバドス、ボスニア&ヘルツェゴビア、カイマン諸島、クロアチア、クラクァオ、コンゴ、ギニア、ジョージア、ガーナ、グアデローフ、香港、ケニア、コソボ、リトアニア,マヨッテ、モンテネグロ、北マケドニア、ポーランド、セルビア、ルワンダ、セントルチア、スロベニア、南アフリカ、スリランカ、トリニダートトバコ、
「B.1.1.7」のデータはこちら「B.1.1.7」
B.変異箇所は?
ウイルスゲノムに23の変異があるが、うち17に重要な変異があり、そのうち、スパイク蛋白質に8の変異がある。
以下表が変異箇所であり、最も注目すべき変異は、「N501Y」である。
N501Y変異ー受容体結合ドメイン(RBD)感染力の増大化とウイルス量の増加に寄与している。免疫系に認識されにくくする特性を持つ。なお、この変異は、南アフリカ変異株でも見られている。
HV 69/70欠失ースパイクタンパク質のコンフォメーション変化(タンパク質の立体配座の形が変わる)につながる可能性がある。
P681HーS1 / S2フューリン切断部位(685と686との間)の近くでの変異である。
ORF8終止コドン(Q27stop)ーORF8の変異であるが、変異による機能変化は不明である。スパイクタンパク質の外側のORF8Q27stop変異。

なお、上記表に掲載されていない「E484K」変異株が、「D.その他」記載の通り、2021年2月1日に追加された。
これは、南アフリカ変異株にもブラジル変異株にも見られた変異である。
C.変異によるウイルス特性の変化は?
A. N501Y変異は、スパイク蛋白質の受容体結合ドメイン(RBD)に位置しており、スパイクRBDとACE2受容体との相互作用により感染力増強(感染速度が速くなる可能性)とウイルス量の増加をもたらすとされる。
B. P681H変異は、感染下細胞が新しくスパイクタンパク質をより効率的に作ることができる変異とされている。
C. 3つの欠失(位置は69/70)は、スパイク蛋白質のコンフォメーション変化(タンパク質の立体配座の形が安定的に変わる)に寄与しており、抗体回避に役立つ可能性のある変異といわれている。
以上の変異により、総じて従来ウイルスより50〜70%速く拡散する可能性を持つ。(他変異株より56%伝染性が高い。との数字も出ている。感染性が約1.5倍高いとも)
子供に感染の可能性が高いとの説もある。
また、実効再生産数R値が+0.4上昇する可能性がある。(R値が0.4から0.7の間)
免疫回避の可能性についての検証は不十分だが、ワクチンの有効性に懸念が生じることもありえる。
なお、下記「D.その他」に記載の通り、近時になり、さらなる変異「E484K変異」があらわれ、これは、抗体反応をすり抜ける「エスケープ変異」ではないか?との一層の懸念が強まっている。
なお、近時発表のロンドン大学衛生熱帯医学大学院のニコラス・G・デイビーズらの研究論文では、これまで最大1.7倍と推定されていた変異株「B.1.1.7」の感染力は、従来型より1.43~1.9倍強いとの推定を発表した。
D.その他
2021年2月1日にイギリスのPublic Health Englandは、英B.1.1.7系統新型コロナウイルスに新たな変異が現れていると発表した。
E484K変異についての報告書 「Investigation of novel SARS-CoV-2 variant Variant of Concern 202012/01 Technical briefing 5」の中で、
「2021年1月26日に、英B.1.1.7系統新型コロナウイルス変異株に、新たに、E484K変異の検出がされた。」としている。
新たに見つかった「E484K変異」は、免疫回避に寄与する変異であるとみなされている。
E484K変異は、南アフリカ由来変異株「B.1.351」系統の「501.V2」にも、ブラジル由来「P.1変異株」にもみられている。
初期の報告段階では、変異体が病気の重症度やワクチンの有効性に影響を与えることを示唆する証拠はなかった。
なお、イギリス政府の科学諮問グループNervtag報告(SAGE meeting paper)によると、「B.1.1.7」は、データは限られているが、死亡リスクが1.3倍に増加とのことだが、詳しい検証はまだ得られていない。
英国ジョンソン首相は、この報告を受け、「英国新型コロナウイルス変異体「B117」が従来株より致命的な証拠がある」と語ったことで、米CDCが、その発言の裏づけの資料を求めている。
米CDCのモデリングでは2021年3月迄に英変異株が主流になるとの見通しを立てている。
なお、デンマークの血清研究所調査研究によると、新型コロナウイルス英国変異株「B.1.1.7」感染者は入院するリスクが高いことがわかった。
B.1.1.7感染者の2,155人のうち、128人が入院し、他の亜種に感染した人より入院する率は64%高いとのことである。。
カナダは、新型コロナウイルス「B.1.1.7」変異株制御に失敗した場合のシミュレーションを示している。
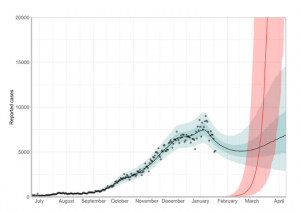
下記図のように2021年3月より急カーブで上昇のシミュレーションとなっている。
(1)-2.「B.1.525」系統
A.いつ、どこで発見されたのか?
イギリス(2020/12/15). ナイジェリア(2020/12/29). フランス(2021/01/06). ガーナ(2021/01/06). ヨルダン(2021/01/07). アメリカ(2021/01/10).デンマーク(2021/01/11). カナダ(2021/01/13).
オーストラリア(2021/01/17).ベルギー(2021/01/19). フィンランド(2021/01/20).
スペイン(2021/01/25). シンガポール(2021/01/31).
「ブリストル・バリアント」とも呼ばれる。
「B.1.525」のデータはこちら「B.1.525 」
B.変異箇所は?
非構造蛋白質orf1ab領域
L4715F
スパイク蛋白質
Q52R
E484K
Q677H
F888L
エンベロープ
L21F I82T
欠失
11288:9
21765:6
28278:3
C.変異によるウイルス特性の変化は?
E484K変異ー抗体をすり抜ける「エスケープ変異」。すでに同様の変異は、英国変異株「B.1.1.7」、南アフリカ変異株「B.1.351」、ブラジル変異株「P.1」に見られる。
Q677H変異ーアメリカ中西部由来変異株、テキサス由来変異株、オハイオ由来株「COH.20G」にも見られる。伝達性を高める変異。抗体をする抜ける『エスケープ変異』との説もある。
新しい変異箇所「E484K」を含んでいる新型コロナウイルス変異株「B1525」については、「以前の感染やワクチンによって獲得された免疫を回避し、より伝染性、致死的になる可能性がある」との懸念が広まっている。
このE484K変異を伴うB117変異株(ブリストル変異株)は、アメリカでも発見されている。
英国の科学者たちは、個の変異株がワクチンの有効性を低下させる可能性があることへの懸念をすでに表明している。
(2)南アフリカ由来変異株
「B.1.351」系統の「20H / 501Y.V2」
A.いつ、どこで発見されたのか?
2020年10月8日に、南アフリカの東ケープ州ネルソンマンデラベイで最初に検出。
2020年12月下旬にザンビアでも確認。
2021年1月25日現在で、感染が報告された国は、26カ国ある。
南アフリカ(2020/10/08)、モザンビーク(11/01)、中国(12/08)、英国(12/12)、スイス(12/14)、ケニア(12/15)、ボツワナ(12/17)、フィンランド(12/19)、日本(12/19)、ドイツ(12/21)、オーストラリア(12/22)、オランダ(12/22)、台湾(12/24)、アラブ首長国連邦(12/26)、韓国(12/26)、ザンビア(12/30)、フランス(12/31)、ノルウェイ(12/31)、スウェーデン(01/03)、オーストリア(01/04)、マヨット(01/07)、カナダ(01/08)、アイルランド(01/08)、ニュージーランド(01/10)、イスラエル(01/10)、ベルギー(01/13)、ガンビア(01/14)、デンマーク(1/16)、ガーナ(01/18)、タンザニア(01/19)、ポルトガル(1/22)、スペイン(1/22)、パナマ(01/23)、アメリカ(01/27)、ヴェトナム(01/31)、ルクセンブルグ(02/02)、トルコ(02/03)、
その他
バングラディッシュ、コスタリカ、コンゴ、イタリア、ルクセンブルグ、ルワンダ、シンガポール、スロベニア、タイ、
など。
俗称「Covid VSOP」とも呼ばれている。
「B.1.351」のデータはこちら「B.1.351 」
B.変異箇所は?
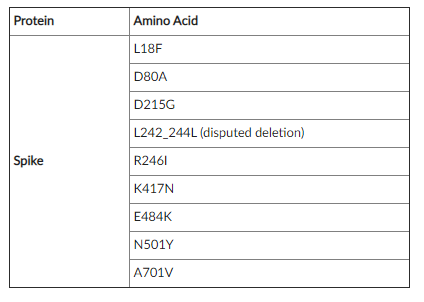
下記表のとおり。
スパイクタンパク質に21個の変異がある。
スパイクタンパク質での主な変異箇所は
D80A
D215G
K417N
A701V
N501Y
E484K
このうち、三変異箇所(K417N 、E484K、N501Y)は、ブラジルの変異株「B.1.1.28.P1」の変異箇所と同じである。
その他のスパイクタンパク質以外の変異箇所としては
E(エンベローブ):P71L
N(ヌクレオチド):T205I
orf1b欠失 11288から11296
がある。
C.変異によるウイルス特性の変化は?
重要な3つの変異(N501Y変異、E484K変異、K417N変異)によるウイルス特性変化(感染の増加とウイルス量の増加に関連か?)に懸念がもたれている。
すなわち
N501Y変異ー感染性が高い原因となる。体内ウイルス量を多くする。これは英国変異株にもある変異。
E484K変異ー免疫応答回避の特性をもち、中和抗体の生成を10分の1低下させる可能性がある。
K417N変異ーE484K変異同様、現在のワクチンでの誘導抗体が亜種に結合し中和しない懸念がある。
これら二つの変異(E484K変異とK417N変異)はいずれもスパイクタンパク質での変異である。
一方、現在開発接種中の世界の主力ワクチンはいずれもスパイクタンパク質を標的としている。
これら2つの変異によって、主力ワクチンの有効性が左右されるのではないか?ワクチン誘導抗体が変異種に結合し中和しない可能性があるのではないか? との懸念を、専門家は持っている。
特に、E484K変異が、一部のポリクローナル抗体およびモノクローナル抗体による中和に影響を与える可能性があることを示すいくつかの証拠が出ている。
なお、これらの変異のうち、「N501Y変異」と「E484K変異」は、英国変異株「B.1.1.7」が有する変異(特に「E484K変異」は、今回新たに加わった変異である。)でもある。
また、「N501Y変異」と「E484K変異」と「K417N変異」は、ブラジル変異株「P.1」にも見られる変異である。
一方、英国変異株が有する変異である「69/70欠失」は、南アフリカ変異株には含まれていない。
現在、この変異が病気の重症度に影響を与えることを示唆する証拠はない。
D.その他
南アフリカで発見の新型コロナウイルスの変異株「501Y.V2」について専門家は下記の懸念を持っている。
①若者に対する感染しやすさがあるのでは?
②再感染の懸念があるのでは?
③ヒト→ヒト感染が容易なのではないか?
④開発ワクチンに対する耐性が、やや高いのではないか?
これについて、昨年末南アフリカ新型コロナウイルス変異株「501Y.V2」特定のNGS-SA等科学者達は501Y.V2特性究明のためのコンソーシアムを結成し、当該変異株に対する当面の課題として以下の点を上げた。
①ワクチン接種者の血液で、ワクチン誘導抗体の効果を検証
②501Y.V2に対してワクチンが機能しない可能性を追及
また、E484K変異についても、一部のポリクローナル抗体およびモノクローナル抗体による中和に影響を与える可能性があるのではないか?との懸念か示されている。
(3)ブラジル由来変異株
総括
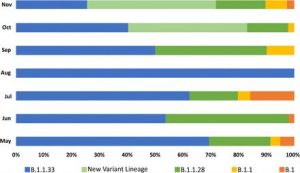
この一年での、ブラジルでの新型コロナウイルス変異株の移り変わりは、激しかった。
2020年5月時点では「B.1.1.33」変異株が主流だった。
「B.1.1.33」変異は、2020年8月には、ブラジル・コロナ変異株のすべてを占めたが、9月以降は、減少に転じた。
「B.1.1.28」変異は、昨年5月から7月まではあったが、8月にいったん淘汰され、9月からマナウスを中心にして「B.1.1.28.P1」変異に分岐され、さらに「B.1.1.28」変異は、12月にはリオデジャネイロを中心にして、「B.1.1.28.P2」に分岐された。
「B.1.1.28.P1」も「B.1.1.28.P2」も、クレード分類上は「B.1.1.28」系統である(一部に「B.1.2.248系統」とされているのは誤りである。)が、現在のクレード分類システムでは、クレード名が3つのサブレベルを超えることができないために。「B.1.1.28.1」または「B.1.1.28.2」というクレード名指定を与えることが出来ないため、「B.1.1.28.P1」あるいは「B.1.1.28.P2」と命名したものである。(P.1 P.2 は「PANGO lineages」に登録済)
現在、ブラジルでは、下記の新型コロナウイルス変異株が循環している。
①2002年5月からすでにある「B.1.1.33」「 B.1.1.28」
②2020年10月頃からある「B.1.1.28.P1」
③2020年12月にリオデジャネイロの研究者によって発見された「B.1.1.28.P2」
なお、P1とP2の名前はパンゴリンに登録されている。
ブラジルでの変異株の変遷は下記図の通りである。
(3)-1 「P.1」(B.1.1.28.P1,)系統
「P.1, 20J/501Y.V3」(ブラジル&日本由来変異株)
A.いつ、どこで発見されたのか?
経緯は複雑である。
昨年(2020年)3月13日に英国へ旅行した女性の感染があった。
後に再感染となるマナウス在住の29歳の女性は、3月3月16日に発症したが、この時のウイルスは、「B.1」であった。
参考サイト「SARS-CoV-2 reinfection by the new Variant of Concern (VOC) P.1 in Amazonas, Brazil」
5月には、サンプルの大部分が系統B.1.1.33であった。
9月までにB.1.1.28が大幅に広がってきた。
10月と11月には「B.1.1.28」が一層優勢になってきた。
2020年10月の時点では、人口の約75%が新型コロナウイルスに感染していると推定された。
このことで、集団免疫状態がすでに確立されたと思われた。
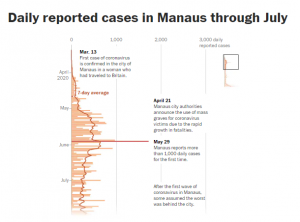
ところが、2020年12月、マナウスで、新変異株が発見された。
「B.1.1.28」系統とみられたが、クレード分類上、3つのサブレベルを超えることができないため、「B.1.1.28.1」変異株とは分類されず「P.1」変異株と名付けられた。
「P.1」変異株の前身とみられる「B.1.1.28」系統変異株には、マナウスの3月から11月のサンプルには新しい系統は見られなかった。
12月にはいり、「B1.1.28.P.1」変異株が急速に優勢となり、マナウスで採取されたサンプルの42%に「B1.1.28.P.1」が確認される。
ちなみに、2020年12月時点での、マナウスから採取されたウイルスの系統別シェアは下記の通りであった。
「B1.1.28」65%-P.1系統を含む
「B.1.161」16%
「B.1.1.33」11%
「B.1.149」5%
「B.1」3%
フィオクルス研究財団研究者によると、12月にコロナウイルス患者から採取されたサンプルの51パーセントに新変異体が存在しており、1月13日までにそれは91パーセントに達したという。
ここで、新事態が発生した。
2021年1月2日、日本の羽田空港で行われていた定期的なスクリーニング中に、ブラジル・マナウス帰りの4人の旅行客から、「P.1」変異株に似た変異株が発見された。
1月9日、日本はWHOに系統B.1.1.28の新しい変異体について通知、当初の日本側の報告では系統B.1.1.248内の変異ということであった。
スパイクタンパク質に12の変異があり、VOC 202012/01および501Y.V2に共通する3変異(K417N / T、E484K、N501Y)が見られた。
2021年1月10日、日本の国立感染症研究所(NIID)は、新しい変異体には12の変異があり、そのうちの1つ(「N501Y」変異のことか?)は英国と南アフリカで見つかった変異体にも存在すると、ブラジルの保健省に伝達した。
当初、日本の国立感染症研究所は、この変異株を「B.1.1.248」系統に属する、と判断したが、ブラジル側の判断では、「B.1.1.28」系統に属するとした。
PANGO lineages分類では「B.1.1.248はB.1.1.28に再割り当てされた」としている。
同時に、日本の国立感染症研究所(NIID)は、「この変異株は、ブラジルの「P.1変異株」に似ているが、一部シーケンスに違いがみられる」とも報告した。

2021年1月12日、ブラジル政府は、「N501YおよびE484K変異を持つP.1として識別された新しい変異株は、12月15〜23日にマナウスで収集された31の陽性PCRテストサンプルのうち13で検出された」と発表した。
この変異は、3月から11月の間に見られたゲノム監視サンプルには存在していなかったとしている。
同日、ブラジル国営のオズワルドクルス財団(フィオクルス)は、アマゾナス州に由来するコロナウイルスの新しい変異体の同定と流通を確認した。
現在、P.1変異株が発見されている国は、下記のとおりである。
日本(2021/01/06)、ブラジル(01/14)、韓国(01/18、フェロー諸島(01/18)、ペルー(01/20)、ドイツ(01/22)、英国(01/23)、イタリア(01/25)、アメリカ・ミネソタ州(01/25)、
その他
スイス、ベルギー、コロンビア、フランス、オランダ、仏領ギアナ、トルコ、アイルランド、スペイン、メキシコ、
「P.1」のデータはこちら「P.1」
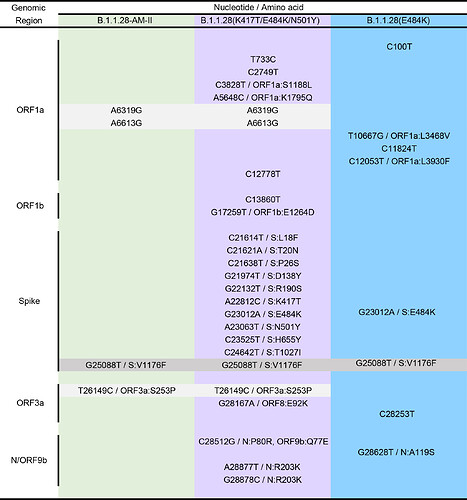
B.変異箇所は?
下記表のとおり。
スパイクタンパク質の変異のうち、三変異(N501Y、E484K、K417N)は、南アフリカの変異株「B.1.351」の変異とほぼ同じ
その他の変異は、
L18F, T20N, P26S, D138Y, R190S, H655Y, T1027I
スパイクタンパク質の外側ORF1b欠失。
欠失部分は
ORF1ab:F681L, I760T, S1188L, K1795Q, del3675–3677 SGF, E5662D0352
ORF3a: C174G
ORF8: E92K
ORF9: Q77E
ORF14: V49L N: P80R
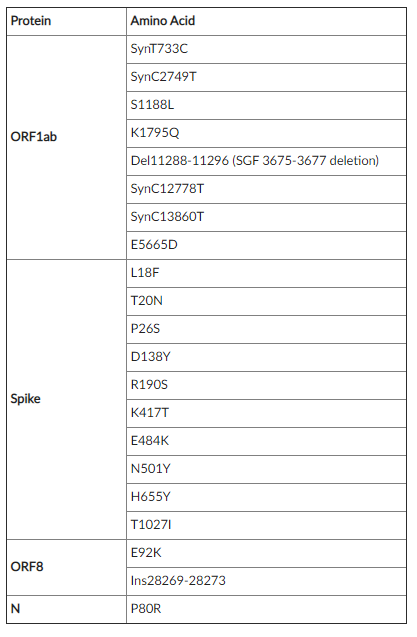
なお、ブラジルのマナウスで発見されたP.1変異株と、日本の羽田空港で発見されたP.1株とのシーケンスの違いは、下記図の紫色部分のみ、日本のシーケンスがブラジルのシーケンスと異なっている部分である。

下記表は、左から「在来のB.1.1.28」「日本・羽田で検出のB.1.1.28変異株」「E484K 変異を有するB.1.1.28変異株」
共通する変異は「薄い灰色」部分。
参考
C.変異によるウイルス特性の変化は?
三変異(N501Y、E484K、K417N)は「B.1.351」と同じ特性変化をもたらしうる。
伝染性または病原性への影響は不明
再感染の可能性や免疫回避の可能性についても、検証が必要。
より伝染性にし、既存の免疫またはワクチン誘発免疫の有効性を低下させる可能性はありうる。
三変異(N501Y、E484K、K417N)のうち、N501Y、E484Kの二変異について、英国変異株、南アフリカ変異株とも同じ変異であり、417については、英国変異株と南アフリカ変異株とが「K417N」であるのに対して、ブラジルP.1変異株は「K417T」の変異である。
変異が、英国、南アフリカ、ブラジルの三地域において、独立して変異していたとすれば、ウイルス側からすれば、同じ選択圧のもとに変異したともいえそうだ。
なお、この変異株の中和効果は大幅に低く、抗体の効力が1〜3倍低下することが証明されている。
再感染がこの特性に基づくものであるかどうかは、まだ明らかにされていないが、Biorxiv公開(査読未了)新型コロナウイルスのブラジル変異株「P.1」の研究「Levels of SARS-CoV-2 lineage P.1 neutralizat」では
「①P.1変異株は、中和抗体からエスケープできる能力を備えている。②再感染の可能性は最初の感染後または、ワクチン接種後もありうる。」
としている。
また、サイト「Brazil Covid variant could cause reinfection in 61 per cent of people」では、
①これまでのブラジル循環株よりも1.4〜2.2倍感染性が高い。
②従来株に感染した人の防御抗体の 25~61%を突破し再感染
③現行のワクチンのP.1変異株に対する有効性はある程度は保たれている。
としている。
(3)-2.「P.2」(別名 B.1.1.28.P2)系統
「20B/S.484K」
A.いつ、どこで発見されたのか?
2020年11月下旬に、リオデジャネイロで検出。
以降、ブラジル全土で検出。
B.変異箇所は?
P.1変異株がスパイクタンパク質に持つ主要三変異(N501Y、E484K、K417N)のうちの一つ、E484K変異を持つ。
C.変異によるウイルス特性の変化は?
E484K変異は、より伝染性にし、以前の感染やワクチン接種により形成された抗体による反応を弱め、免疫力低下につながる可能性がある。
P1バリアントと同様、P2が何らかの変化をもたらすかどうかはまだ明らかでない。
D.その他
P.1かP.2かの2つのブラジルの変異体のいずれかによる感染が見つかった場合、市の公衆衛生サービス(GGD)が調査を開始することになっている。
(3)-3 「P.3」(B.1.1.28.3)系統(「VUI-21MAR-02(P3)」)
A.いつ、どこで発見されたのか?
フィリピンの中部ビサヤ(Visayas)地方の
ダバオ(Davao)地方から3症例
カラバルソン(Calabarzon)から2症例
バリクバヤン(在外フィリピン人)から8症例
その他6症例
英国のハーリンゲイ・ロンドン地域とウェストミッドランズでも三例発見された。
B.変異箇所は?
E484K,
N501Y,
P681H,
P1176F
141-143del.
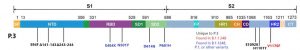
C.変異によるウイルス特性の変化は?
まだわからない。
D.その他
日本で、同様に変異株がフィリピンからの旅行客から検出されている。
3月12日、日本の国立感染症研究所は2月25日にフィリピンから成田空港に入国した60代男性から変異株が検出と報告。
N501Y変異とE484K変異を有していた。
(3)-4 「VOI N.9」 (B.1.1.33)系統
A.いつ、どこで発見されたのか?
B.変異箇所は?
NSP3:A1711V、
NSP6:F36L、
S:E484K、
NS7b:E33A
C.変異によるウイルス特性の変化は?
まだ解明されていない。
昨年8月まで「B.1.1.33」はブラジルで循環していた主力株であり、その時点での変異箇所は「N:T29148C N:T27299C」であった。
今回発見の変異株は2020年8月に登場し、南東部、南部、北部、北東部などのブラジル各州に広がったものとおもわれる。
参考
(4)アメリカ由来変異株
(4)-1.オハイオ由来変異株
「COH.20G / 501Y」 と「COH.20G/677H」
A.いつ、どこで発見されたのか?
2020年12月下旬オハイオ州コロンバスで発見。
以降、他の米国州でも検出。
B.変異箇所は?
「COH.20G / 501Y」はスパイク蛋白質にのみに「N501Y」変異。
「COH.20G / 677H」は、スパイクタンパク質とMおよびNに変異。
C.変異によるウイルス特性の変化は?
伝達性、病原性、免疫回避の変化の証拠はまだない。
(4)-2.テキサス由来変異株
「20C-US」
A.いつ、どこで発見されたのか?
5月下旬にテキサス州で発見。
B.変異箇所は?
スパイクタンパク質Q677H変異
C.変異によるウイルス特性の変化は?
フューリン切断部位に隣接するスパイクタンパク質Q677H変異であることから、伝達性に影響するのでは?との懸念がある。
D.その他
日本では、2020年11月、慶應義塾大学医学部チームは、全国13の病院の患者からのサンプル関東の中に関東地方の患者に「20C」変異株を発見している。
参考
(4)-3.カリフォルニア由来変異株
「Cal.20C」または「B.1.429」または「20C/S:452R;/B1429」
A.いつ、どこで発見されたのか?
2020年3月にデンマークで発見
2020年5月にカリフォルニア州で発見。
2020年7月に米ロサンゼルス郡で確認。
2020年11月から12月にかけて、北カリフォルニアの症例における変異頻度は3%から25%に上昇。
2021年1月、カリフォルニアの複数の郡で頻度多く発見。
2020年11月下旬から12月上旬間ではテストサンプルの約3.8%、12月下旬から2021年1月上旬にかけては25.2%以上、と、急速に増加している。
最初3月にデンマークで確認、
なお、オレゴン州立大学とオレゴン健康科学大学は、新型コロナウイルスのカリフォルニア変異株(L452R変異)が、2020年3月には検出されていたとしている。
カリフォルニアには、現在、B.1.427およびB.1.429の二種類の変異株が流通している。
B.変異箇所は?
5つの変異がある。
ORF1a:I4205V、
ORF1b:D1183Y、
S:
S13I。
W152C
L452R
後者の3つの変異(プラス、従来からある変異のD614G)は、スパイクタンパク質に影響を及ぼし、細胞への結合を容易にする可能性がある。
特にL452R(別名B1429)変異に注目。
うち1つは、英国のとは異なる変異であることに注目。
参考「Emerging SARS-CoV-2 Variants」
C.変異によるウイルス特性の変化は?
L452R変異は、S1部分(1–685)での変異であり、細胞表面受容体認識を担う部分での変異であるところから、中和抗体の影響を受けにくくなる可能性はありうる。
カリフォルニアでのいくつかの大発生に関連しているとみられるが、確証は得られていない。
ワクチンの有効性への影響についても調査中。
JAMAのレポート
「SARS-CoV-2 Viral Variants—Tackling a Moving Target」では、
①新型コロナウイルスの変異株は進化的選択のプロセスを通じ優勢に
②L452R変異はモノクローナル抗体結合に影響を与える可能性
③近時の変異株は急速に抗原変異傾向を示している
としている。
(4)-4.米中西部由来変異株
A.いつ、どこで発見されたのか?
オハイオ州で12月と1月に発見。中西部の複数の州でも発見。
「中西部変異株」とも呼ばれる。
B.変異箇所は?
スパイクタンパク質のQ677H変異、
Mタンパク質のA85S、
およびヌクレオカプシドタンパク質のD377Y
C.変異によるウイルス特性の変化は?
伝達性、病原性、免疫回避の変化の証拠は、まだない。
(4)-5.中南部&南西部由来「Q677P」変異株
A.いつ、どこで発見されたのか?
2020年12月24日にバージニア州で発見。
以降、ワシントン、ニュー目メキシコ、カンザス、フロリダ、テキサス、イリノイ、スカロライナ。カリフォルニア、マサチューセッツ などで発見
参考「COVID3D」
B.変異箇所は?
「Q677P」
S1 / S2フューリン切断部位が「685と686との間」であり、この切断部位に近い部位での変異( Q677P置換)なので注目されている。
C.変異によるウイルス特性の変化は?
「中和抗体からのエスケープ変異」なのではないのか?との見方もある。
さらに、アメリカで発見の7つの変異株のいずれも、「677」のポジションに変異をきたしていることについて、専門家は、収斂進化の表れではないかと、危惧する向きもあるようだ。
この変異株は2020年8月中旬まで検出されていない。
2021年2月3日時点で、米国からGISAIDに寄託の102,462ゲノムのうち2,327以上を占めているとされている。
(4)-6.ニューヨーク由来「B.1.526」変異株
A.いつ、どこで発見されたのか?
2020年11月にニューヨーク市で収集されたサンプルに現れ始めた。
他の国への感染拡大はほとんどない模様。
参考「A Novel SARS-CoV-2 Variant of Concern, B.1.526, Identified in New York」
「B.1.526」のデータはこちら「B.1.526」
B.変異箇所は?
L5F
T95I
D253G
S477N
E484K
D614G
A701V
![]()
C.変異によるウイルス特性の変化は?
抗体をすり抜ける「エスケープE484K変異」があり、ワクチン効果を弱めるのでは?との危惧が広がっている。
南アフリカ変異株と共通特性を持ち、より伝染性が高く制御が困難。
E484K変異は抗体エスケープ変異特性をもち、S477N変異はヒト細胞へのバインディング強化の特性を持つ。
(5)ナイジェリア由来変異株
A.いつ、どこで発見されたのか?
8月3日と9月9日収集サンプルのうち、ナイジェリアのオスン州の2人の患者から発見された。
B.変異箇所は?
スパイクタンパク質にP681Hの変異をもつ。
「B.1.1.7」との共通変異があるかどうかは、まだ確認されていない。
C.変異によるウイルス特性の変化は?
変異箇所(681)が、フューリン切断部位(685と686との間)に直接隣接していることで、ACE2受容体との親和性が増すのではないのか?という点に懸念がもたれている。
(6)デンマーク由来株
「クラスター5」
A.いつ、どこで発見されたのか?
デンマークでは、ミンクの新型コロナウイルスの感染があり、そこに5つのクラスターが存在していた。
これを「クラスター5」とよんだ。
ここには、スパイクたんぱく質にいくつかの特異な変異があった。
2020年11月2日までに検出した変異体には、人間に感染する懸念のある変異があった。
ユトランド半島北部にヒト感染と思われる症例が12あり、いずれも抗体反応を弱化させる変異が見られた。
B.変異箇所は?
69–70deltaHV
Y453F
I692V
M1229I
S1147L
C.変異によるウイルス特性の変化は?
クラスター5株のさらなるヒトの症例は検出されなかった。
クラスター5の亜種はもはや人間に循環していないとの一応の評価の段階にとどまっているが、英国変異株との関係など、なお、注視が必要とされている。
参考「Cluster 5」
(7)インド変異株
(7)-1.「N440K」変異(インド南部由来株)
インド南部で「N440K」変異蔓延。
これら変異株は免疫回避E484K変異と、より高い伝達率をもつN501Y変異を有しているという。
インドCCMB(細胞分子生物学センター)の科学者Divya Tej Sowpati氏によると、今話題になっている新型コロナウイルス変異株「N440K」(B.1.36)(下記↓図ピンク色)は、今年の2月まで一定の感染拡大をしていたが、3月に、急速に衰え、今は変異株のほとんどが「B.1.617」変異株に置き換わったという。
参考
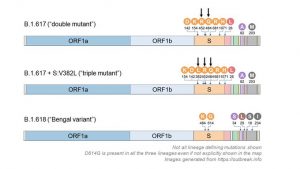
(7)-2.二重変異株(B.1.617)
変異箇所 E484K、L452
(7)-3.三重変異株(B.1.617+スパイク蛋白質V382L)(B.1.617)
変異箇所 E484K、L452、V382L
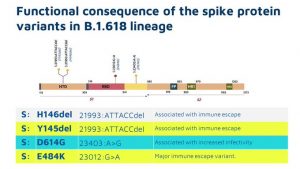
(7)-4.ベンガル変異株(B.1.618)
変異箇所 E484K、D614G 欠失がH146とY145

(8)その他の変異
A.「Fin-796H」
フィンランドで変異株『Fin-796H』を発見。
ワクチンに対する感染性と潜在的耐性については、まだ、詳細はわかっていない。
B.「B.1.1.316」
2021年2月日本で発見、感染由来不明
「B.1.1.316」には
①「E484K」変異(免疫回避特性)はあるが
②「N501Y」変異(感染性が高い特性)はない。
なお「B.1.1.316」変異株は日本だけの変異株ではなくアメリカでも発見されている。
参考
「新型コロナウイルスSARS-CoV-2 Spikeタンパク質 E484K変異を有するB.1.1.316系統の国内流入(2021年2月2日現在)」
C.ブラジルでの「B.1.1.33」系統が「E484K」変異をもち再登場
変異箇所
NSP3:A1711V、
NSP6:F36L、
S:E484K、
NS7b:E33A
D.「アンティグア・コビッド」変異株
カリブ海にあるアンティグアに旅行した人から発見
イングランド南東部で2例発見
3月4日にVUI(調査中変異株)に指定
スパイク変異E484KとN501Yを有す
参考「New Antigua Covid-19 strain declared as ‘Variant Under Investigation’ – what we know so far」
E.フランスのブルターニュ地方で発見の変異株「ブルターニュ変異株」(Breton variant)
「levariantbreton」とも「variant du Trégor」とも「20C 」とも呼ばれる。
詳細はまだわかっていない。
参考
「欧州疾病予防管理センターにおいて、2021年5月4日時点で懸念されているSARS-CoV-2変異株一覧」
参考サイト「SARS-CoV-2 variants of concern as of 4 May 2021」
懸念される変異株(VOC)
-300x95.png)
関心のある変異株(VOI)
-300x164.png)
監視中の変異株
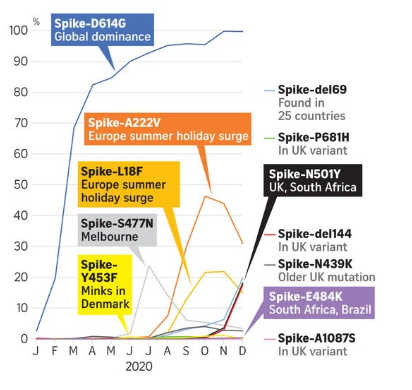
3.懸念される変異はなにか?
以上、現下、世界に蔓延しつつある新型コロナウイルスの変異株の現状についてみてきた。
現在の世界の新型コロナウイルスの症例は、日本にも見られるように、漸減傾向にあると同時進行的に、世界で変異株が優勢増加している事態をどう見るのか?、専門家は「変異株による本格的な波が近づいているのではないのか?」と、逆に危惧しているという。
またも専門家は「パンデミックが発生してから1年以上経った今、より伝染性の高い新型コロナウイルス変異株が出現している理由は何なのか?」ということにも疑問を持っている。
いくつかの懸念が専門家の間から上がりつつある
すなわち
①ワクチンが標的としているスパイクタンパク質での変異が多い。
②抗体を回避する変異がある。
③再感染への懸念や風土病化への懸念やADE(抗体依存性感染増強)への懸念がある。
④死亡率の上昇が懸念される変異がある。
⑤通常のプライマー・セットでは、変異株はPCR検査では見つけられない
⑦若年層への感染強化の懸念がある。
などである。
さらには、二つの異なる新型コロナウイルスの変異株同士が再結合しての、ハイブリッド組み換え変異株も誕生している。
再結合は英国由来変異株「B.1.1.7」と米カリフォルニア由来変異株「B.1.429」で、組換え体はロスアラモス国立研究所Bette Korberが発見した。
更なる組換えもありうるとしている。
(1)懸念される共通の変異
最大の懸念は、英国変異株、南アフリカ変異株、ブラジル変異株共通の変異が、相互の関係なしに生まれてきていることである。
新型コロナウイルスが複製を続けていくうえで、必然的に迫られた変異が、世界で、時と場所を変えて、同じ選択のもとに、同じ変異を迫られているとも解釈される。
以下は、その代表的な共通変異である。
A.「N501Y変異」ー英国変異株、南アフリカ変異株、ブラジル変異株P.1 とP.2 、それにフィリピンで発見のP.3、米・ オハイオ変異株「COH.20G / 501Y」に共通の変異。
受容体結合ドメイン(RBD)感染力の増大化とウイルス量の増加に寄与している
B.「E484K変異」ー英国変異株(2021年1月に変異判明)、南アフリカ変異株とブラジル変異株P.1 とP.2 、それにフィリピンで発見のP.3 に共通の変異。
ワクチンの有効性を低下させる「エスケープ変異」である。
免疫力低下。一部のポリクローナル抗体およびモノクローナル抗体による中和に影響を与える可能性があることを示すいくつかの証拠がある。
ロンドン大学ユニバーシティカレッジのフランソワバロウ教授は「E484K変異には抗体認識を低下させる特性がある。 そのため南アフリカの新型コロナウイルス変異種が以前の感染またはワクチン接種によって確立された免疫保護を回避させてしまうことになってしまう」と述べている。
C.「K417N変異」ー南アフリカ変異株、ブラジル変異株P.1の場合は、変異は「K417T」であった。
E484K変異同様、現在のワクチンでの誘導抗体が亜種に結合し中和しない懸念がある。
上記三変異のハブとなりえるのが、南アフリカ変異株「501.V2」である。
ロンドン大学のフランソワバロウ教授は「南アフリカ変異株「501.V2」は、抗体認識を低下させる変異を有している」との認識である。
その他の共通変異としては次のものがある。
D. 「L18F変異」ー南アフリカ変異株とブラジルP.1.変異株と共通。変異はコンフォメーション(蛋白質の立体構造) 変化をもたらし、ウイルスのACE2受容体へのバインディング力を強くする。
E.「欠失(69~70)」(del H69/V70)ーデンマーク変異株(クラスター5)と英国変異株(B.1.1.7)と共通。スパイクタンパク質のコンフォメーション変化(タンパク質の立体配座の形が変わる)により、需要委への結束力を強くする可能性がある。
F.「D614G変異」ー武漢株から最初に変異し、その後、続く各変異株においても、その変異を維持し続けている。
これらの主要変異による特性変化をまとめてみれば下記のようになりうる。
1.S蛋白質RBD変化で親和性増加→感染性促進
N501Y
参考
「N501Y COVID-19 Variants — Should We Be Worried?」
「Effect of mutations in the South African SARS-CoV-2 variant on transmission and neutralization」
2.ウイルス極性変化で抗体の認識低下
E484K
3.界面塩橋破壊でRDBのACE2結合親和性低下
K417N
参考
「Impact of South African 501.V2 Variant on SARS-CoV-2 Spike Infectivity
and Neutralization: A Structure-based Computational Assessment」
4.S蛋白質立体構造安定化
D614G
5.選択圧からの選択と脱出
欠失(69~70)
参考
「Natural deletions in the SARS-CoV-2 spike glycoprotein drive antibody escape」
ニューデリーのINYAS主催プレゼンテーションで、ジャミール氏が、二つの変異(N501YとE484K)を持つウイルス特性につき講演し、「同じウイルスで501と484の両変異同時発生の場合、抗体反応回避か、抗体効果低下の可能性あり」と発言。
また「484変異は、アミノ酸の極性変化(Polarization)をもたらす」とも講演したのが注目されている。

(2)免疫応答回避メカニズムはあるのか?
ウイルスの中に、免役応答回避メカニズムを持っているものは、ほかにもある。
例えば、畜産の世界で今恐怖となっているアフリカ豚熱ウイルス(ASFV)は、その典型的なウイルスである。
このウイルスの免疫応答回避メカニズムについては、昨年、私のブログ記事「加速するアフリカ豚コレラ(ASF、アフリカ豚熱)ワクチン開発ー絶望から光明へ転換はあるか?」で紹介した。
このウイルスの場合の免疫応答回避メカニズム(immune evasion mechanisms)は、免役応答能のアップ・レギュレートとダウン・レギュレートの二つをうまく使い分けて、自然免疫応答および適応免疫応答の両方の回避をおこなっている。
新型コロナウイルスの場合は、次の免疫回避メカニズムが働いているといわれている。
「Covid-19: Perspectives on Innate Immune Evasion」によると
「ACE2受容体はインターフェロン誘導遺伝子(ISG)であり、IFNの上方・下方制御に関与している。」
として
コロナの自然免疫回避は下記の3つによって行われている。としている。
①受容体の認識から回避
②インターフェロンの転写を阻害
③I 型インターフェロンのシグナル伝達を阻害
この免疫回避メカニズムを、変異が、助長するのかどうか?が問題である。
新型コロナウイルスの変異株が、抗体反応を回避するかどうか?につき、現在、いくつかの研究成果がある。
すなわち
①南アフリカ変異株感染回復者からの血漿につき検証した研究
「SARS-CoV-2 501Y.V2 escapes neutralization by South African COVID-19 donor plasma」
②免疫血清の中和力価を検証した研究
「Neutralization of SARS-CoV-2 lineage B.1.1.7 pseudovirus by BNT162b2 vaccine-elicited human sera」
③mRNAワクチン誘発抗体についての検証研究
「mRNA vaccine-elicited antibodies to SARS-CoV-2 and circulating variants」
④invitroでコロナ回復者から採取の血漿とコロナウイルスを共培養。
血漿は7継代ウイルスを完全中和したが、73日目にE484K置換、80日目に中和に耐性のある変異。E484K変異が抗体反応を回避
「SARS-CoV-2 escape in vitro from a highly neutralizing COVID-19 convalescent plasma」
などである。
新型コロナウイルスの免疫回避メカニズムに関わっていると疑われているのが、次の変異株である。
①南アフリカ変異株「B.1.351」(2020年10月)
E484KとK417Nの組み合わせ
②ブラジル&日本変異株「P.1(B.1.1.28.1)」(2021年1月)
E484KとK417T変異
③カリフォルニア変異株「B.1.429」(2020年7月)
L452R変異
これらの変異株のうち。特に注目されているのが、「E484K変異」と「K417T変異」である。
E484K変異(科学者たちの間では「Eek」とも呼ばれている。)が、地域と時期に関わらず、離れた世界の地域で、一斉に招じたということは、「この中和エスケープをもたらす特定の変異がウイルス進化の一般的な解決策として機能する可能性がある」との研究もあるようだ。
まさに「変異株は動く標的」であり、抗原の変動によつて、中和への有効性が低下している事態といえる。
(3)変異によるウイルスのコンフォメーション変化(蛋白質の立体構造変化)はあるか?
どうして、新型コロナウイルスの変異株が、感染が早く拡がるのか?
これについては「変異によるコンフォメーション(ウイルスの立体構造)の変化で結合力が強まるため」との専門家の見解が強い。
「E484K変異」「K417T変異」両変異によるコンフォメーション(タンパク質の立体構造)変化が、ACE2結合へ影響を与えているのではないか?との研究がいくつかある。
E484K変異がスパイクRBD-ACE2親和性を増強し、E484K、K417N、N501Y変異(501Y.V2変異体)の組み合わせがN501Y変異体単独よりも大きなコンフォメーション(タンパク質の立体構造)変化を誘発し、エスケープ変異体をもたらす可能性がある、としている研究もある。
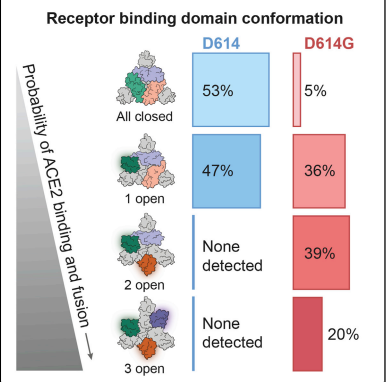
スパイク蛋白質は、受容体結合ドメイン(RBD)の上下の位置に応じて、受容体にアクセスできない(閉じた)状態になることも、アクセスできる(開いた)状態になることもある。
変異によるコンフォメーション(タンパク質の立体構造)の変化によって、開いた、アクセスしやすい状値になることによって、受容体への結合がしやすくなるということである。
上記の図でいえば、AとBの状態とでは、ACE2に対する結合可能性のあるタンパク質のポジションが異なるということである。
なお、コンフォメーション(タンパク質の立体構造)の変化でオープンになるほど、なぜ感染性が強まるのか?については、マサチューセッツ大学ウースター校医学部のJeremy Luban氏の説によれば、変異によってプロトマー間の相互作用が邪魔されることによって、コンフォメーション(タンパク質の立体構造)がオープンになればなるほど、受容体へのバインディング結合力が強くなる(ウイルスの感染力が高まる)、とのことである。
下記図は「D614G変異」での例である。
(4)再感染の可能性はあるのか?
米ネバダ州在住の男性が新型コロナウイルスに再感染した事例が英医学誌「ランセット」で報告された。
初回感染から再感染までの時間的経過は下記の図の通りである。
2020年3月25日発症→4月18日PCR陽性→5月26日陰性→5月28日再発症→6月5日PCR陽性
ゲノム解析が十分ではなかったが、「2084欠失、6018挿入」のみ確認されている。
再感染の可能性として、上記研究では、次の三つの可能性を上げている。
A.二回目の感染は、第一回目の感染に比し、非常に高用量のウイルス感染によって引き起こされ、重篤化した。
B.再感染は、より毒性の高い、またはより毒性の高い変異株によって引き起こされた。
C.抗体依存性増強のメカニズムが原因だった可能性もある。
この例での再感染がどの変異株によってもたらされたかは、ここの現場でのゲノム解析が不十分だったために、検証されていない。
また、初感染から再感染までの期間があまりに短いため、果たして、初感染での抗体生成が十分だったのか?との疑念も残る。
E484K変異を有する。 ブラジル変異株に感染のバイア州北東部サルバドール市45歳女性も「再感染」を起こしていたが、この場合は、 初感染5月20日、再感染10月26日と間に5ヶ月もの差があった。
ブラジルのマナウスでは、いったん市民の大半が感染した後、変異株P.1の登場で、地域的に再感染に見舞われた地域もある。
英国公衆衛生サービス(PHE)の科学者による予備調査結果によると
①新型コロナウイルスに感染した人は、すくなくとも、5か月間は免疫を有している可能性
②しかし、抗体を有している人も、ウイルス感染能力をなお有している可能性がある
③再感染からの保護率83パーセントである。
との研究成果もある。
また、ブラジルのマナウスでの再感染については、四つの要因があったとしている研究もある
すなわち
①第一次感染での発症率が過大に評価されていた。
②第一次感染により形成の免疫抗体は、第二次感染時に、既に衰え始めていた。
③第二次感染時におけるウイルス(特にP.2変異株)の中に存在し下E484K変異が、免疫回避の特性を持っていた。
④第二次感染時のP.1系統ウイルスは、第一次感染時ウイルスよりも感染力が強かった。
これらの4要因が複合化し、集団免疫状態を崩したとしている。
なお、EricFeigl-Ding博士によると、以前に新型コロナウイルスに感染した人でも、南アフリカB1.3.51変異株への再感染では、初感染の人同様に重篤な疾患に対し保護されていない、とのことである。
(5)風土病化への懸念、ADEへの懸念はないのか?
ウイルスの風土病化(Chronic)は、野生動物や家畜を宿主とするウイルスにも、よく見られる現象である。
家畜を宿主にしてのウイルスに見られる風土病化・慢性化では、潜伏期間が急性に比して極端に長く、また、ウイルス排出期間も長期にわたることなのが゛特徴であった。
臨床症状もことなっていた。
ヒト宿主の新型コロナウイルスの風土病化についても、十分な注視が必要である。
新型コロナウイルスは風土病化するのか?という懸念については、次の見解もある。
すなわち
通常のウイルスの場合は、最初の感染後、ヒト適応免疫システムは、メモリーBリンパ球、およびメモリーTリンパ球を含む一連の防御機構が発達しうるのだが、これが、新型コロナウイルスにも適用できるのか?に疑問が残るとしている。
ここでポイントとなりうるのが「抗原連続変異」(antigenic drift)が起きうるのかどうか?なのだという。
免疫系によって認識されるウイルスゲノムの突然変異の無作為な蓄積によりウイルスの抗原性が著しく変化し、免疫系による攻撃からの回避を助けることがあるという現象である。
一方、再感染で、初回感染よりも重症化したとの例も多く出ているが、これが、初回感染によりできた(または、ワクチン接種により形成された不十分な抗体が導引となっての抗体依存性増強現象によるものなのか?は、これからも十分な検証が必要である。
その他、VAERD(ワクチン関連増強呼吸器疾患)、VAH(ワクチン過敏反応)についての知見には、いまだ未解明のものが多い。
なお「抗体依存性増強現象」については、私のブログ記事「新型コロナ ウイルス ワクチン 接種と「抗体依存性感染増強」(ADE)現象誘発の懸念」」も、ご参照。
(6)変異による死亡率(fatality rates )の上昇の懸念はあるのか?
さきにも記述したが、2021年1月21日に、イギリス政府の科学諮問グループによるNervtag報告(SAGE meeting paper)「B.1.1.7の重症度に関するNERVTAGノート」で、「データは限られているが、「B.1.1.7」変異株は、死亡リスクが1.3倍に増加」と報告され。英国ジョンソン首相は、この報告を受け、「英国新型コロナウイルス変異体「B117」が従来株より致命的な証拠がある」と語ったことで、波紋が世界中に広がった。
米CDCが、その発言の裏づけの資料を英国政府に求め、。米CDCのモデリングでは2021年3月迄に英変異株が主流になり、変異ウイルスの感染率は従来株よりも約50%高い、との見通しを立てた。
そして、「B.1.1.7変異株のコミュニティ感染はすでに全米で進行中」ともした。
研究報告によると、B.1.1.7は、2020年後半から米国の州間で広がっている可能性があり、これからの数週間、早急かつ果断な公衆衛生措置が取られない限り、この変異株の持つ高い伝搬率と、その結果としての「実効再生産数(R)」の上昇によって、米国における新型コロナウイルスの死亡率と罹患率に、壊滅的な結果をもたらす可能性がある、としている。
また、ロンドン大学衛生熱帯医学大学院Kucharski氏は「感染力が従来株より50%高い新型コロナウイルス変異株は致死性の高い変異株よりはるかに大きな問題になりうる。 「実効再生産数(R)」率が1.1で死亡率0.8の変異株は一か月死者数129人。 死亡率50%増加で死者数193人となりうる。 一方、感染力が50%高い変異株では死者数978人になりうる。」と警告している。
1月中旬、イギリスの二つの大学の研究グループが、別々に英国のNERVTAGに研究論文を提出した。
それによると、変異株の死亡リスクが約30%増加しているとした。
エクセター大学と英国公衆衛生サービスによる他の研究でも、死亡者数が多いことがわかった。
また、3月15日にネイチャー発表の論文
「Increased mortality in community-tested cases of SARS-CoV-2 lineage B.1.1.7」によると
新型コロナウイルス英国変異株「B.1.1.7」は、従来株に比して死亡危険性が61%(42〜82%)高くなると推定された。
なぜ新型コロナウイルス変異株がリスキーか?
伝送速度が高い変異株は、死亡率が高い変異株より危険になりうるからだ。
致死率が50%増加すると、50%多くの死亡が発生はする。
しかし、伝達率が50%増加すると、指数関数的に増加し、わずか2、3か月で25倍の感染例が発生するからだ。
(7)若年層への感染強化への懸念はないのか?
インペリアル・カレッジ・ロンドンの研究者によると、20歳未満の人々は新型コロナウイルスの英国変異株「B.1.1.7」感染症例に高い割合を占める可能性が高いことが分かった。
研究は「封鎖が実施されていたが学校が開いていた時期」に注目し、この期間での20代未満の世代の感受性、またはより明白な症状についての調査をしたところ、20代未満世代に有意の影響を見て取れたという。
このことについて、政府諮問グループ(NERVTAG)のニールファーガソン教授は、英国での2回目のロックダウン中、5週間または6週間の封鎖期間中に学校だけは開いていたが、その間に、新型コロナウイルスの変異株に子供が感染する率が有意にに多かったとしている。
また、米国小児医師たちは新型コロナウイルスの変異株が、子供たちの間に感染しやすい状況にあるのではないか?と懸念している。
すでに270万人近くの子供たちが陽性となっており、直近2週間で、子供たち377,000近くの新症例があり、16%増加しているという。
特に「B.1.1.7.」変異株は警戒としている。
なお、WHOが新型コロナウイルス変異株が子供に及ぼす影響を検証することにした。
WHOのKerkhove博士は
①症状は年少の子供ほど軽度になる傾向にあり、総じて無症候性が多い。
②英国変異株は幼児を含むすべての年齢層で伝染性が増加している。
変異株が特に子供を標的にしている証拠はない。
との見解のようである。
(8)ワクチン効力の無力化はあるか?
ワクチンの有効性を無にする変異
現在の世界の新型コロナウイルス対策は、ウイルス側の「変異株の拡大」と、ヒト側の「社会的ディスタンス遵守とワクチン接種率の拡大」との競争状態にある。
その意味で、ワクチンの有効性を低下させうる変異が、ヒト側にとって一番の脅威となりうる。
特に脅威なのは「B.1.1.7」変異株の出現であり、CDCのディレクター ロシェル・ワレンスキー博士は、伝染性の高い新型コロナウイルス変異株(特にB.1.1.7株の出現)は、パンデミック統制を逆転させ、この病気に対する「私たちのすべての努力を弱体化させる」可能性があるとさえ述べている。
また、米マサチューセッツ総合病院研究者たちの研究「Vaccine-induced antibodies may be less effective against several new SARS-CoV-2 variants」
では、ワクチンによって誘導され生成された抗体はいくつかの新型コロナウイルス変異株に対しては、効果が少ない。としている。
変異に対する各ワクチンの有効性は?
これまで開発を終え上市された代表的な新型コロナウイルスワクチンのメーカーが発表している、ワクチンの有効性の数値は下記のとおりである。
ファイザー/ BioNTechー95%
モデルナー94.5%
アストラゼネカー70%
ジョンソン&ジョンソンー66%以上、重度から重篤な症例の予防には85%有効
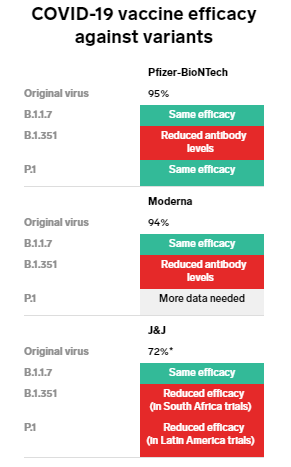
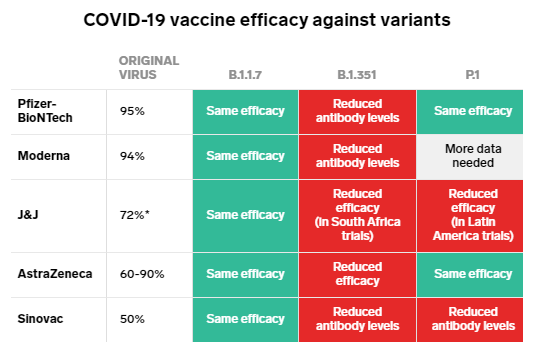
参考「One chart shows how well COVID-19 vaccines work against the 3 most worrisome coronavirus variants」
どの変異がワクチンの有効性を損なっているのか?
変異株に対して、これらの有効性がどう変わっていくかが問題だが、総じて、南アフリカ変異株に対する各ワクチンの有効性は、目減りしているというのが、現在の状況のようである。
ファイザー&BioNTechの新型コロナウイルスワクチンが南アフリカ変異株に対しては低い有効性を示していることは、すでに研究成果に現れている。
NEJM発表のテキサス大学研究では、3つの組換えウイルスを用いた実験で南ア「B.1.351」への有効性低下を証明している。
二つのワクチン(NovavaxとJohnson&Johnson)の治験結果が、南アフリカ変異株に対しワクチン効果が少なかったことで、新型コロナウイルスが「ワクチン回避変異」( vaccine-escape mutations)してるのではないのか?との疑念を関係者に与えている
Novavaxワクチンは英国で実施の臨床試験では89%有効だったが、南アフリカでは49%に低下した。
また、ジョンソン&ジョンソンのワクチンの効果は、米国で72%効果的であったのに対し 、ラテンアメリカでは66%、南アフリカでは57%であった。
「E484K変異」はワクチン共通の敵
今回、英国新型コロナウイルス変異株「B.1.1.7」に新たに発見された「E484K変異」がワクチン効果をどれだけ弱めるのか? について、専門家の関心があるようだ。
2021年2月1日、英国公衆衛生庁(PHE)は報告書を公表し、この中で、2021年1月21日に、英国変異株に「E484K」変異が見られるとした。
「E484K」変異は、抗体をすり抜ける「エスケープ変異」であることから、にわかに、現在開発・接種中のワクチンの効力に、関係者の懸念が生じてきた。
中和抗体はウイルスが細胞に侵入するのを防ぐはずだが、受容体RBDの変化により、これらの抗体がE484K変異を持つ変異体を認識しにくくなる可能性がある。
また、この変異を持つ変異株は、多くの人々がすでにCOVID-19に感染している集団であっても、より容易に広がる可能性があり、以前の抗体を持つ人々の再感染を可能にしうる。
英国研究者は、現在、3つのB.1.1.7変異が免疫反応にどのように影響したかを検証中である。
すなわち、これら検証では、ファイザーBioNTechワクチンBNT162b2の初回投与から3週間後に患者の免疫反応をテストするとともに、変異株に対するワクチンの効果検証も始めている。
マサチューセッツ工科大学(MIT)と米国のハーバード大学の国際的な研究者チームが、ファイザーワクチンとモデルナワクチンの有効性を検証するため、新型コロナウイルスの10種類の株のスパイクタンパク質を中和する能力について分析をした。
検証対象は
①中国の武漢株②デンマークのB.1.1.7、③B.1.1.298、④B.1.429、⑤P.1、⑥P.2、⑦B.1.351、およびコウモリコロナウイルスWIV1-CoV、
等であった。
その結果、
ファイザーワクチンの中和力は、野生株の中和力よりも
①B.1.1.7に対しては43%に減少
②B.1.1.298に対しては29%に減少
③B.1.429に対しては34%に減少
した。
さらに、2021年3月4日にネイチャーメディシンで発表されたワシントン大学医学部の研究 によると 新型コロナウイルスに感染したか、ワクチン接種を受け、防御抗体がある人々は、変異株から保護されない可能性があるのとの懸念を示している。
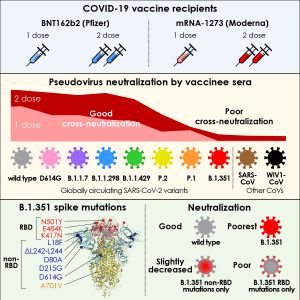
どのワクチンも南アフリカ変異株に対しては弱い
南アフリカ変異株に対しては、すでに、二つのワクチン(NovavaxとJohnson&Johnson)の治験結果があり、南アフリカ変異株に対しワクチン効果が少なかったとしている。
また、南アフリカの疫学者Abdool Karim氏は
「新型コロナワクチンが南アフリカの変異株の「501Y.V2」(または、 B.1.351)に対して有効であるという証拠はまだない」と指摘した。
関係者は、南アフリカ変異株が「ワクチン回避変異」( vaccine-escape mutations)してるのではないのか?との疑念を持ち始めている。
「Circulating SARS-CoV-2 variants escape neutralization by vaccine-induced humoral immunity 」では、ファイザーのワクチンの中和力について、新型コロナウイルス野生株と変異株(「B.1.1.7」「B.1.1.298」「B.1.429」「P.1」「B.1.351」)に対する比較を行っている。
ファイザーのワクチンが、南アフリカ変異株(B.1.351)に対しては、効果が薄い(2/3)との立証をしたNEJMの研究論文もある。
さらに、2月18日のワシントンポストでは
「ファイザー、モデルナ両ワクチンは南アフリカ新型コロナウイルス変異株への有効性を低下」との記事を載せ、その中で、Lindsey R. Baden博士らの研究では、既存のコロナ株に対する有効性は証明されたが、南アフリカ変異株に対してはブースターショットまたは最新ワクチン開発措置が必要との研究結果が出たとしている。
変異に対応しての既開発ワクチンの設計変更は可能か?
「新型コロナウイルスの変異株に備えて、現行のワクチンの設計変更はされるのか?」については、
「インフルエンザワクチンの例をモデルにすべき。WHOとの毎年の新しいワクチンレシピに基づき変更となるのでは?」との専門家の意見もある。
スタンフォード大学ブリッシュ博士は「新型コロナウイルス変異株とワクチン接種とのスピード競争が始まってる」とし、ファイザーとモデルナは既に変異株対応を始めているし、アストラゼネカは、南ア変異株に対応できないたことを認め、秋までワクチン組成微調整を目指す、という。
国立アレルギー感染症研究所のファウチ博士は、ジョンソン&ジョンソンの新型コロナウイルスワクチンの治験結果が南アフリカ変異株に対し低いワクチン効果を示したことについて「ワクチン製造業者は必要に応じワクチンを再処方するため「容易に調整できるように機敏」でなければならないと述べた。
また、感染拡大を続ける新型コロナウイルス変異株「B.1.1.7」に対し、ファウチ博士は、「二つの強力なツールがある」とし、「それは①B.1.351変異株対応ワクチンの開発 ②マスクと手洗いの公衆衛生習慣の継続であるとし、「それには7月までに国民全員接種を終えることが必要」としている。
現時点では、変異株への現行ワクチン有効性に疑問符が付いており、
サイト「One of the strongest weapons against COVID-19 variants may be a vaccine the FDA hasn’t approved yet」に見るように、「現行開発ワクチンで最も変異株対応ができるのは、これからFDA許可までの間変異株対応設計変更を一早くできうるアストラゼネカだけではないのか?との極端な見方さえあるようだ。
変異対応のための既存ワクチン開発会社の試み
他のワクチン開発会社も、変異株に対する自社ワクチンの有効性を高めるための試行錯誤をし始めているようである。
新型コロナウイルスの変異株に対し、ワクチンメーカーはどのように対応するのか?
「How would COVID-19 vaccine makers adapt to variants?」によると、
①ファイザーとモデルナは
遺伝子コードを交換し変異株遺伝子コードに適合させる。
②アストラゼネカは
変異スパイク遺伝子を使用し無害なインフルウイルスを増殖させる。
とのことであり、②は、時間がかかるとのことである。
③Novavaxは
南ア変異株に対し第2b相試験開始
④VBIVは
VBIとCEPIが共同で「B.1.351」対応ワクチン開発で提携した。
CEPI(感染症流行対策イノベーション連合)は、VBI社(VBI Vaccines Inc)が開発する新型コロナウイルス変異株対応方ワクチン「eVLP」開発支援を決定。
eVLPワクチンは、ウイルスのエンベロープをターゲットにしたワクチンで変異株に対応できるとしている。
ブースターショットの試みも
ホワイトハウス上級顧問アンディ・スラビット氏は
「ワクチンメーカーはブースターショットで新型コロナウイルス変異株防御力強化できるか検討開始」と述べた。
ModernaとPfizer / BioNTechは変異株対応のためワクチン更新を計画。必要に応じ、ブースターショットも計画している。
モデルナは、さらに変異株に対し効果的なブースターショットを計画している。
新変異株と南アフリカ変異株二つのブースターショットを実験。 ワクチン接種済の人やサルの血液を使った研究では、英国変異株は中和抗体産生減少はなかったが、南アフリカ変異株には抗体産生が6分の1減少を示したとしている。
変異株対応ブースターショットの「第2相試験+患者への投与」を開始した。
その他の有力ワクチンメーカーであるファイザー、ジョンソン&ジョンソンも、変異株に対応したワクチン強化のためのブースターショットやその他の取り組みの開発に着手しているとされている。
変異株にあわせたワクチン設計変更や微修正の試みも
また。アストラゼネカとパートナーのオックスフォード大学は、利用可能な新しいバリアントに合わせた微調整バージョンを、秋までに用意することを目指している。
既存ワクチンの新型コロナウイルス変異株への微調整が難しいのは、設計変更が可能なほど、十分にモーフィングしたかどうか?( morphed enough)をどの段階で判断するか?
(モーフィング( Morphing)とは、ウイルスの「突然変異への変形」という意味。)
つまり、変異の過程にあるのか?最終変異の段階に落ち着いたのか?の見極めがむづかしぃということのようだ。
ちなみに、ワシントン大学保健指標評価研究所の最新のCOVID-19予測によると、
①最悪のシナリオは 南アフリカ変異株「B.1.351」によるパンデミックで米国で5月1日までに約654,000人死亡
②ワクチン接種キャンペーンによっても、集団免疫効果が得られない場合 ワクチンの有効性は50%に低下
との最悪シナリオも想定されているようだ。
一回接種か?二回接種か?接種感覚を空けるのか?
ファウチ博士は、2021年1月25日の「World Economic Forum」の席で、「二回の接種を求められているワクチン接種の一回目と二回目の接種間隔が所定の間隔よりもあいてしまうと、ウイルスに抗体回避できる変異「エスケープ・ミュータント」を促進させてしまうかもしれない」と警告した。
現在の二回接種を条件にした各ワクチンにおける一回目と二回目との間隔は下記のとおりである。
ファイザー&BioNTechー21日
モデルナー28日
アストラゼネカー4週から12週以上
ウェイルコーネル医科大学のジョンムーア教授によると、「突然変異の発生を可能にする自然選択圧力は、不完全な免疫反応から来ており、これがエスケープ変異を起こす原因となる。
もうひとつは、決められた二回のワクチン接種の1回目と2回目の投与の間の、抗体が弱く、エスケープ変異体を作成できる期間に発生するリスクがある」としている。
米-疾病対策センターのアドバイザーは、「新型コロナウイルスワクチンであるmRNAワクチンの2回目の接種を遅らせると、患者の変異株に対する防御が弱くなる可能性がある」と警告。
「2回目接種を遅らせることを正当化するには、十分な証拠がない。」とした。
また、一回目と二回目との接種間隔をあけることは、ウイルスの変異を加速させるのでは?ウイルスのワクチン耐性を高めるのでは?との懸念をする専門家もある。
一方で、新型コロナウイルスワクチン接種を一回にとの論議がイギリスなどで高まっている。
イギリスではセカンドショット最大12週間延期を決定した。
専門家は2回目8週間遅れ迄は影響なしとしている。
ただ一回目接種抗体がいつまで持続か?は不明ともいっている。
一回目接種により形成された中途半端な抗体形成はウイルスに変異を促しADE現象誘発リスクもあるとの専門家の意見もある。
3月9日サイエンス発表マギル大学&プリンストン大学研究
「Epidemiological and evolutionary considerations of SARS-CoV-2 vaccine dosing regimes」によると
二回接種条件の新型コロナウイルスワクチン接種の二回目接種時期を遅らせると抗原連続変異の可能性ありとしている。
第二世代ワクチン開発の試みも既に始まった
新しい考え方によるワクチン開発の動きもすでに出てきた。
GlaxoとCureVacは、1つの新型コロナワクチンで複数の変異体に対処する第二世代ワクチンをmRNA技術により開発を計画しているとのことである。
また、リバプール大学のヒスコックス氏は
「Sタンパク質ベースのワクチンが最初に登場したが、次のラウンドではヌクレオカプシド(N)タンパク質標的追加が必要」と、第二世代ワクチンへの見通しを述べている。
ただ、これらの第二世代のま新型コロナウイルスワクチンの展望があったとしても、フランスのCOVID-19科学評議会のメンバーたちは、「新型コロナウイルスワクチンの登場があっても、パンデミックの終焉は、変異株に対し有効なワクチンが世界中に公平に配布された場合にのみ可能」との展望を示している。
なお、モノクローナル抗体については、Regeneron 社のモノクローナル抗体イムデビマブ(REGN10987)とカシリビマブ(REGN10933)で構成されるREGEN-COV(カシリビマブおよびイムデビマブ抗体カクテル)が、英国変異株(B.1.1。 7)南アフリカ変異株(B.1.351)に対しても、有効であることを確認している。
その一方で、別の研究では
「Regeneron 社のモノクローナル抗体REGN10933は、B.1.1.7に対し中和活性を維持しているが、B.1.351および「クラスター5」スパイクタンパク質に対しては、中和活性力を失っている」ことを示した研究結果もある。
さらに、韓国のSKバイオサイエンスは、CEPIの支援を受けて、新しい新型コロナウイルスワクチン開発に進出。変異株に対応できる万能型ワクチン「GBP510」開発&上市をする。とのことである。
(9)変異株を検出する検査体制の強化はできるのか?
メルボルン大学ドハーティ感染免疫研究所パーセル教授は
「英変異株が英国で発見されたのは、英国が他の国よりも多くの新型コロナウイルスのゲノム配列を研究していたために外ならない。」という。
またインド医師会副会長のラジーブ・ジャヤデヴァン氏も、「ゲノムサーベイランス能力を持つ国ほど多くの新型コロナウイルス変異株を報告」とし、「最近の亜種を最初に報告した国は英国であり、一番優れている国だ」といっている。
ゲノム・シーケンスにより変異株摘出体制に優れた国こそ、新型コロナウイルス対策の先陣をきりうる、との見方だ。
現在のPCR検査体制では、変異株の検出がスムーズにできない。
そのため、変異株発見の公表が、遅れてしまう。
ちなみに、英国のケント・バリアントといわれる変異株は、変異株自体が発見されたのは昨年9月だったが、重要変異があることが分かったのは、昨年12月だった。
この変異株発見に至る十分の時間的余裕で、公表されるころには、この変異株は、既に世界循環していた。
まして、変異によって、無症状患者が多く出ていては、なおさらのことである。
米FDAは「南アフリカの新型コロナウイルス変異種は、PCR検査も、偽陰性ですりぬける怖れ」という警告を、今年の1月初めに出している。
アメリカでも変異株を見つけるには従来にない方法を確立しなければならないとし、変異株の存在をキャッチできる「プライマー」開発を始めている。
この場合、感度と特異性との、二つのバランスをとることが必要としている。
日本政府は「新型コロナウイルス変異株を短時間で検出しうる新たな検査体制を3月から敷く」というが、これは「これまで塩基配列のわかっている変異株に焦点を当てた、PCRのプライマリー設計変更を行う」ということなのだろう。
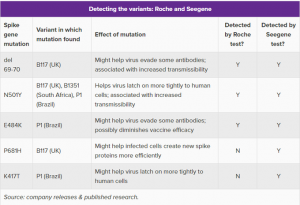
ここにきて、フランスの検査機器メーカーのNovacyt社は「SNPsig SARS-CoV-2VOC識別キット」を世に出し、これまでのPCR検査機器そのままでも、変異株を検出できる検査キットを、このたび、発売した。
その他の検査機器メーカーも、同様の検査機器を開発している。
その先陣を切っているのが、Rocheと韓国のSeegeneの両社である。
ロッシュはコバスSars-Cov-2バリアントセット1(「Cobas®SARS-CoV-2 Variant Set1)テスト
これは、現在分析ラボや病院で使われている「Cobas6800 / 8800」マシンで使用できるように設計されている。
Seegeneは「Allplex Sars-Cov-2 Masterアッセイ」
両者の性能の比較は下記↓のとおり
おわりにー大陸を横断しての新型コロナウイルスの同時変異は「収斂進化」の帰結なのか?ー
以上、現時点(2021年2月8日時点)での、新型コロナウイルスの変異状況と、現時点でのその影響を、ざっと見てきた。
気になるのは、「はじめに」にも書いた「3つの異なる変異(N501Y変異とE484K変異とK417変異)が、3つの異なる大陸で、異なるLineageで独立して出現したにもかかわらず、同じ突然変異((N501Y, K417N、E484K)が、Sタンパク質のRBDで同じ変異を共有している事実は、偶然ではなく必然の「収斂進化」( convergent evolution)の帰結なのではないのか?」という疑問である。
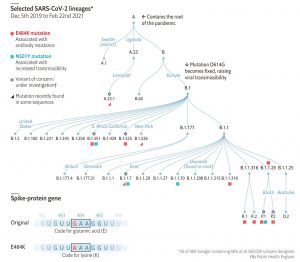
これら三変異がウイルスに進化上の利点をもたらした可能性がある。
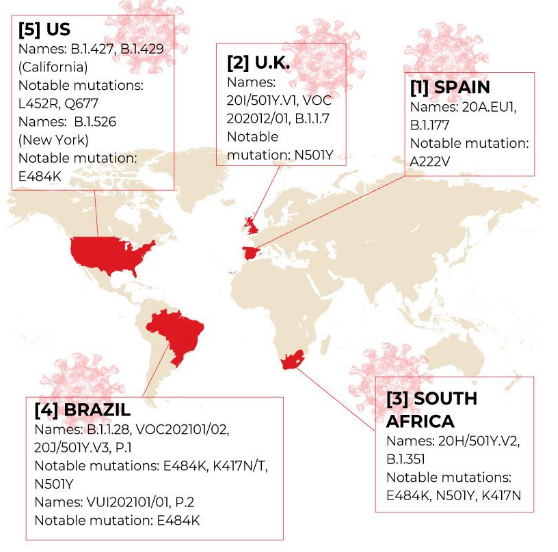
日本における変異株への感染者数は
3月5日現在
埼玉40、兵庫37、新潟31、神奈川15、東京14、大阪12、京都9、静岡7、福島5、鹿児島5、岐阜4、群馬3、岡山3、山梨2、滋賀2、茨城1、千葉1、栃木1、長野1、石川1、空港検疫57 合計251例
となっている。
しかし、実態はこの数字ではわからない。
2021年2月に入り、日本の感染研究所は、感染由来の不明な変異株「B.1.1.316」が日本で発見されたと発表した。
変異株状況は、毎日変化する。
そして、それに従って、ワクチンの有効性も、日々変化してきている。
どちらが勝つかという視点よりは、もっと長期的視点にたっての対策も、準備しておくべきなのだろう。
「変異株による第四波は必ずやってくる」との覚悟を持って。 (終わり)
著者 笹山登生(2021年2月8日記述)
参考文献一覧
参考(0)はじめに
「SARS-CoV-2 spike D614G change enhances replication and transmission」
「Experts in Singapore divided on effect of new Covid-19 strains in virus fight」
「Genetic Variants of SARS-CoV-2—What Do They Mean?」
「B.1.1.7: What We Know About the Novel SARS-CoV-2 Variant」
「Worrisome New Coronavirus Strains Are Emerging. Why Now?」
「SARS-CoV-2 Variant Pseudovirus」
「This Key COVID Mutation Is a Scary Sign of What’s to Come」
「Establishment and lineage dynamics of the SARS-CoV-2 epidemic in the UK」
「SARS-CoV-2 mutations acquired in mink reduce antibody-mediated neutralization」
「Genetic sequencing is vital as vaccine will put pressure on virus to mutate, says Dr V Ravi」
「On the founder effect in COVID-19 outbreaks: how many infected travelers may have started them all? 」
「Evaluating the Effects of SARS-CoV-2 Spike Mutation D614G on Transmissibility and Pathogenicity」
「A Primer on Coronavirus, Variants, Mutation and Evolution」
参考 2.変異株は、いつどこで発見されたのか?変異箇所はどこか?ウイルス特性は何か?
参考 2-(1)-1 「B.1.1.7」系統の「VOC-202012/01」
「Estimated transmissibility and impact of SARS-CoV-2 lineage B.1.1.7 in England」
「What do we know about the new coronavirus variant?」
「Emergence of SARS-CoV-2 B.1.1.7 Lineage — United States, December 29, 2020–January 12, 2021」
「Hospitalization risk 64 per cent higher with B.1.1.7 coronavirus variant: study」
「NERVTAG Presented to SAGE on 21/1/21」
参考 2-(1)-2.「B.1.525」系統
「New Covid-19 variants: what we know about the Kent, South Africa and Brazil strains」
「‘Bristol Covid variant’ that may diminish vaccine efficacy discovered for first time in US 」
参考 2-(2)南アフリカ由来変異株
「What we know about the South African variant of Covid-19」
参考 2-(3)ブラジル由来変異株
総括
「Genomic characterization of a novel SARS-CoV-2 lineage from Rio de Janeiro, Brazil」
「Genomic characterisation of an emergent SARS-CoV-2 lineage in Manaus: preliminary findings」
参考2-(3)-1 「P.1」(B.1.1.28.P.1)系統
「New coronavirus variants could cause more reinfections, require updated vaccines」
「Brasil confirma circulación de nueva variante del coronavirus en el Amazonas」
「SARS-CoV-2 reinfection by the new Variant of Concern (VOC) P.1 in Amazonas, Brazil」
「Brief report: New Variant Strain of SARS-CoV-2 Identified in Travelers from Brazil」
参考 2-(4)-3.カリフォルニア由来変異株
「New COVID variant with 5 mutations identified in California」
「‘California’ COVID-19 variant detected in multiple Oregon counties, U.K. variant pops up in Bend」
「Cremation backlog prompts air-quality rules to be temporarily suspended in Southern California」
「New California Variant May Be Driving Virus Surge There, Study Suggests」
「SARS-CoV-2 (COVID-19) S protein RBD (L452R)」
参考 2-(4)-5.中南部&南西部由来「Q677P」変異株
「Study Identifies Seven New Covid-19 Variants in U.S. Carrying ‘677’ Mutation」
「7 homegrown COVID variants detected in US: study」
「Study Identifies Seven New Covid-19 Variants in U.S. Carrying ‘677’ Mutation」
「Batch of homegrown coronavirus mutations seen in US」
参考 3 懸念される変異はなにか?
「Experts fear complacency with COVID-19 cases in decline, variants on the rise」
「Covid-19: Why more contagious variants are emerging now, more than a year into the pandemic」
「What’s the concern with COVID-19 variants?」
「New coronavirus variant: What do we know?」
「Why more contagious Covid-19 variants are emerging?」
「Two variants have merged into heavily mutated coronavirus」
参考 3-(1)懸念される共通の変異
「South Africa coronavirus variant: What is the risk?」
「Coronavirus variants with two specific mutations may elude immune responses: Expert」
「The same covid-19 mutations are appearing in different places」
「Virus expert warns of “convergent evolution” amid COVID-19 mutations」
「Worrisome New Coronavirus Strains Are Emerging. Why Now?」
参考 3-(2))免疫応答回避メカニズムはあるのか?
「Covid-19: Perspectives on Innate Immune Evasion」
「SARS-CoV-2 mutations allow escape from clinical monoclonal antibodies」
「Covid-19: Perspectives on Innate Immune Evasion」
「How does SARS-CoV-2 evade the immune defences?」
「Structural basis for translational shutdown and immune evasion by the Nsp1 protein of SARS-CoV-2」
「SARS-CoV-2 Viral Variants—Tackling a Moving Target」
「Fast-spreading COVID variant can elude immune responses」
「The Impact of Mutations in SARS-CoV-2 Spike on Viral Infectivity and Antigenicity」
参考 3-(3)変異によるウイルスのコンフォメーション変化(蛋白質の立体構造変化)はあるか?
「The Science Behind Why New Covid Variants Are Spreading Faster」(動画)(英語)
「Structural and Functional Analysis of the D614G SARS-CoV-2 Spike Protein Variant」
「Why the New Covid-19 Variants Could Be More Infectious」
「Conformational transition of SARS-CoV-2 spike glycoprotein between its closed and open states」
「Mutation of Covid-19 detrimental to certain T cell immune responses: Study」
参考 3-(4)再感染の可能性はあるのか?
「Genomic evidence for reinfection with SARS-CoV-2: a case study」
「Resurgence of COVID-19 in Manaus, Brazil, despite high seroprevalence」
「What we know about covid-19 reinfection so far」
「Covid reinfection symptoms: New variant sees sufferers infected again – symptoms to spot」
参考 3-(5)風土病化への懸念、ADEへの懸念はないのか?
「Will SARS-CoV-2 become endemic?」
参考 3-(6)変異による死亡率(fatality rates )の上昇の懸念はあるのか?
「NERVTAG Presented to SAGE on 21/1/21」
「Covid 19 coronavirus: Why new strains are so dangerous and how we can stop them」
「New COVID Strain Spreading Across U.S.: What We Know」
「Why scientists think the UK variant of Covid-19 could be more deadly」
参考 3-(7)若年層への感染強化への懸念はないのか?
「Virus Updates: Variant From South Africa Detected in US; LA Mayor Gets Vaccine」
「COVID-19 Increases in Children; Two Die in Fort Worth」
「What We Know So Far About How The New Covid Variant Impacts Kids」
「The intriguing features of COVID-19 in children and its impact on the pandemic」
参考 3-(8)ワクチン効力の無力化はあるか?
「Major vaccine manufacturers already researching, planning for COVID-19 variant spread」
「New forecasts show possibility of spike in COVID-19 cases and deaths in the spring」
「New clinical trials raise fears the coronavirus is learning how to resist vaccines」
「Scientists express concern about how new Covid-19 variant will respond to vaccine」
「Variants du coronavirus : lequel d’entre eux est le moins neutralisé par le système immunitaire ?」
「Comprehensive mapping of mutations to the SARS-CoV-2 receptor-binding
domain that affect recognition by polyclonal human serum antibodies」
「Understanding The New COVID-19 Variants And Vaccine Effectiveness」
「Safety and Efficacy of the BNT162b2 mRNA Covid-19 Vaccine」
「Efficacy and Safety of the mRNA-1273 SARS-CoV-2 Vaccine」
「Pfizer, Moderna vaccines have reduced effectiveness against South African variant, studies show」
「Circulating SARS-CoV-2 variants escape neutralization by vaccine-induced humoral immunity 」
「Worrisome New Evidence That COVID-19 Vaccines Are Less Effective Against New Coronavirus Variants」
「Some SARS-CoV-2 variants evade mRNA vaccine-induced humoral immune, says study」
「COVID-19 Vaccines Might Be Changed to Fight New Versions of Virus」
「Neutralizing Activity of BNT162b2-Elicited Serum — Preliminary Report」
「The latest on the coronavirus pandemic and vaccines」
「Covid variants could ‘undermine all of our efforts’ if virus spreads globally, CDC director says」
「The Race Against SARS-CoV-2 Variants 」
「Looking beyond COVID-19 vaccine phase 3 trials」
「Investigation of novel SARS-CoV-2 variant Variant of Concern 202012/01」
「UK coronavirus variant has developed a vaccine-evading mutation, government document reports」
「UK coronavirus variant develops vaccine-evading mutation」
「Variants du coronavirus : lequel d’entre eux est le moins neutralisé par le système immunitaire ?」
「Comprehensive mapping of mutations to the SARS-CoV-2 receptor-binding
domain that affect recognition by polyclonal human serum antibodies」
「Tackling Covid-19 variants with next-generation vaccines」
「New COVID-19 forecasts show the possibility of a spring spike in deaths with new variants」
「What do we know about the SARS-CoV-2 vaccines so far?」
「Fauci Says He’s Worried About Delaying Second Vaccine Dose」
「‘Escape Mutations’ May Drive New COVID Resurgence」
[Insights on SARS-CoV-2’s Mutations for Evading Human Antibodies: Sacrifice and Survival]
「How will COVID-19 vaccines adapt to the new COVID-19 variants emerging?」
「Delaying the second COVID-19 shot could mean less protection, CDC says」
「Could too much time between doses drive the coronavirus to outwit vaccines?」
「How would COVID-19 vaccine makers adapt to variants?」
「One Shot Offers COVID-19 Protection, But for How Long?」
「GSK and CureVac Take Aim at COVID-19 Variants with Second-Generation mRNA Vaccine」
「Rise of variants sparks push for all-in-one Covid vaccines」
「SARS-CoV-2 variants and ending the COVID-19 pandemic」
「UK coronavirus variant develops vaccine-evading mutation」
「Tackling Covid-19 variants with next-generation vaccines」
「The Race for Vaccine Against COVID-19 Variants Heats Up; 3 Stocks to Consider」
参考 3-(9)変異株を検出する検査体制の強化はできるのか?
「UK coronavirus variant spreading rapidly around the world likely emerged from a single infection」
「Nations with greater genomic surveillance will report more Covid variants」
「New methods might be needed to catch COVID-19 virus variants in Manitoba」
「Covid 19 Testing qPCR Detection & Extraction Kits」
「COVID-19 test makers are adapting for variants」
参考-おわりにー大陸を横断しての新型コロナウイルスの同時変異は「収斂進化」の帰結なのか?ー
「Emerging SARS-CoV-2 Variants」
「Resurgence of COVID-19 in Manaus, Brazil, despite high seroprevalence」
「A Guide to Emerging SARS-CoV-2 Variants」
「SARS-CoV-2変異株B.1.1.7由来のスパイク変異体コレクション」
「COG-UK report on SARS-CoV-2 Spike mutations of interest in the UK」
「Concerning Variants of SARS-CoV-2」
「Emergence and Evolution of a Prevalent New SARS-CoV-2 Variant in the United States」
「Epidemiological update:Occurrence of variants ofSARS-CoV-2 in the Americas 」
「新型コロナウイルス感染症(変異株)の患者等の発生について」(日本の厚生労働省)
「Insights on SARS-CoV-2’s Mutations for Evading Human Antibodies: Sacrifice and Survival」
今年の秋口になり、秋冷を迎える頃、世界で二つの新型コロナウイルスの変異が、話題となった。
一つは、欧州での第二波または第三波到来の主導力となったとみられる新型コロナウイルスの変異株「20A.EU1」であり、もう一つは、デンマークを中心とする養殖ミンクとその養殖業者に感染が広まった「クラスター5」とよばれる新型コロナウイルスの変異株の存在である。
これまでの新型コロナウイルス変異の変遷
これまでの新型コロナウイルスの変異の変遷は次のとおりである。
ボロニア大学の新型コロナウイルスゲノム解析 によれば
①武漢で2019年12月L株
②最初の変異はS株。2020年初出現
③1月中旬以来V株とG株
④2020年2月末にG株はGRとGH株に変異
となる。
参考「University of Bologna genome analysis: The six strains of SARS-CoV-2」
その後は大きな変異はなく、最適化されてきた。
今年の8月時点では、これらの6株が下記の分布で、世界に併存し続けてきた。
①L株 武漢株、徐々に消失
②S株 2020年初め、米国、スペイン、徐々に消失
③V株
④G株 2020年2月末に分布 GR株とGH株に変異 アジア
⑤GR株 ヨーロッパ、イタリア、南アメリカ、アジア
⑥GH株 フランス、ドイツ、北米、アジア
「D614G変異」に対する世界の懸念
今年の6月12日にアメリカのスクリップス研究所から発表された論文が「D614G変異」について触れ、一挙に、この「D614G変異」が話題を呼んだ。
参考「Mutated coronavirus shows significant boost in infectivity」
この段階で、スクリップス研究所研究はコロナは「D614G変異」で感染力を増しているが、開発ワクチンはこの変異に対応できるとしている。
また、D614G変異が、スパイクの数を増やし、また、スパイクの機能をより安定化させており、これによって、ウイルスの細胞への侵入を容易にさせているともしている。
論文「The D614G mutation in SARS-CoV-2 Spike increases transduction of multiple human cell types」では、変異で感染能力が8 倍増加するともしている。
また、中国の論文では
①コロナウイルスの感染能力は2.4〜7.7 倍に増加
②スパイクたんぱく質表面のウイルス量は4.7 倍に増加
とする研究もある。
参考「新冠重要发现:需警惕!多个研究团队发现,发生D614G突变的新冠病毒,感染人类细胞的能力或提高9倍左右」
新型コロナウイルスの欧州株に見られる「D614G」変異というのは 、フーリン蛋白質開裂部残基配列第614番目において 、 D(アスパラギン酸)がG(グリシン)に変異しているということである。
変異サイトがS1 とS2の中間にあるため、S2への親和性が増すことによる安定性が増強されるとのことである。
参考「Genomic diversity and hotspot mutations in 30,983 SARS-CoV-2 genomes」
新型コロナウイルスのD614G変異の意味 としては
①変異後(SG614)は変異前(S D614)より、効率的にACE2発現細胞に感染
②これは S1の排出が少ない。 S蛋白質の疑似ビリオンへの抱合が多い。 ことによる。
③変異後(S G614)が変異前(S D614)より安定している。
とのようである。
参考「The D614G mutation in the SARS-CoV-2 spike protein reduces S1 shedding and increases infectivity」
上記図のシーケンスにおけるD614G変異箇所は下記
赤字の(D→G)
のところ である。
TGTGVLTESNKKFLPFQQFGRDIADTTDAVRDPQTLEILDITPCSFGGVSVITPGTNT
SNQVAVLYQ(G)VNCTEVPVAIH
ADQLTPTWRVYSTGSNVFQTRAGCLIGAEHVNNSYECDIPIGAGICASYQTQTNSPRRAR
全配列は下記ご参照
参考「Recombinant Human Novel Coronavirus Spike glycoprotein(S) (D614G), partial」の9行目から最後まで
今年の2月までは、世界の新型コロナウイルスには、この変異は見られなかったが、3月頃から変異が見られ始め、5月には70%に変異が見られるようになった。
8月に入り、インドからマレーシアに帰国した男性から発生したクラスター感染者45名の中に、D614G変異株が発見され、改めて、D614G変異に、世間の注目が集まった。
参考「Southeast Asia Detects Mutated Virus Strain Sweeping the World」
G株とD614G変異との関係
先に、現在、世界に流行している新型コロナウイルスには6株(S L V G GR GH)があるとしたが、これと、D614G変異との関係は次のとおりである。
GR=D614G変異とG204R変異
GH=D614G変異とQ57H変異
このように、現在の新型コロナウイルス変異状況は
D614G変異(G型)がさらに細分化され、
GR型(D614G変異プラスG204R変異)
と
GH型(D614G変異プラスQ57H変異)
に分かれ。、その他、
S型(L84S変異)、
V型(L37F変異とG2151V変異)
などがある。
アジアで主流はS型、ヨーロッパで主流はG型とGR型、北米南米で主流はGH型となっている。
変異による新型コロナウイルスの特性変化は下記のとおりである。
①D614G 感染性上昇
②N439K, L452R, A475V, V483A, F490L,Y508H 一部中和抗体への耐性向上
③D614G+I472V 感染性と中和抗体への耐性向上
④N-結合型糖鎖の欠損 特にN331とN343の糖鎖欠失が感染性を弱化。
N234Qは耐性向上
N165Qは感受性向上
参考
「The Impact of Mutations in SARS-CoV-2 Spike on Viral Infectivity and Antigenicity」
「我国科学家又发表一篇Cell论文!详细剖析SARS-CoV-2刺突蛋白突变对病毒感染性和抗原性的影响」
「详细剖析SARS-CoV-2刺突蛋白突变对病毒感染性和抗原性的影响」
ここにきての新しい新型コロナウイルス変異株の登場
①エスカピアンス
9月2日「エスカピアンス」(escapians)と呼ばれる新しい変異株の登場を示唆する論文がヒューストン・メソジストから発表された。
この研究で、James M. Musser氏ら研究者たちは、ワクチンを逃れることができる突然変異体ウイルス(「エスカピアンス」と呼ばれる)または薬物や他の治療法に抵抗することができる突然変異体を生成できる自然発生の突然変異によってウイルスが勢いを増す懸念があると示唆した。
「中和モノクローナル抗体(mAb)CR30022による認識の低下」というのだが、詳細はまだわからない。
参考
「HOUSTON METHODIST COVID-19 STUDY SHOWS RAPID SPREAD AND POTENTIAL FOR MUTANT VIRUSES」
②「20A.EU1」という新型コロナウイルス変異株が、欧州を席巻
10月25日、バーゼル大学が、ショッキングな論文を発表した。
参考「Emergence and spread of a SARS-CoV-2 variant through Europe in the summer of 2020」
新型コロナウイルスの変異株「20A.EU1」が、6月にスペイン北東部農業地帯で出現し7月からヨーロッパ中に感染拡大し、現在の欧州各国ロックダウンを招いている。というものである。
6つの特徴的な遺伝子変異を持ち、この変異が免疫応答回避の特性をウイルスに与えているという。
新型コロナウイルスの変異株「20A.EU1」は、 ウイルスのスパイク、 ヌクレオカプシド、 および ORF14タンパク質 のアミノ酸を修飾する変異を特徴とする。
「20A.EU1」のゲノム配列は、 ①スパイクタンパク質の変異A222V や ②核タンパク質のA220V など、 6つ以上の位置で祖先配列とは異なっている。
変異箇所は「A222V」である。スパイクタンパク質シーケンスの222番目のアラニンがバリンに変異している。
この変異株は、交叉免疫が効きにくい点に懸念が持たれている。
「20A.EU1」株の進化型として「20A.EU2」株がある。
変異箇所は「S477N」である。
ヨーロッパ各国別の型は
フランス 20A.EU2
イギリス 20A.EU1
オランダ 20A.EU1
スイス 20A.EU1
ベルギー S98F
スペイン 20A.EU1
ノルウェー20A.EU2
である。
参考「Emergence and spread of a SARS-CoV-2」
変異の進化過程の順序は
D614G
→
20A.EU1
→
20A.EU2
となっている。
参考「Emergence and spread of a SARS-CoV-2 variant through Europe in the summer of 2020」
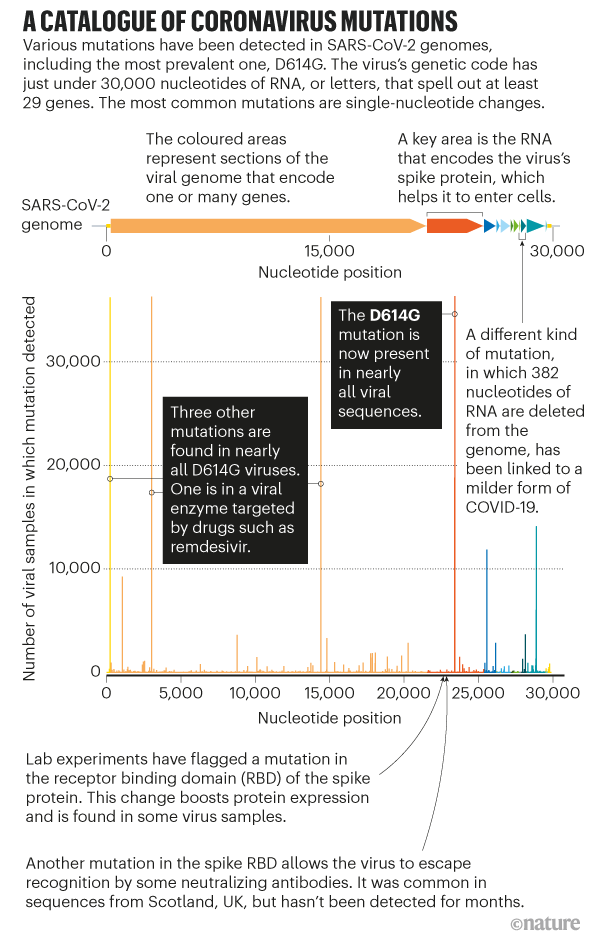
現在、変異株20A.EU1 による感染が見られているのは、欧州12ヵ国(スペイン、イギリス、アイルランド、スイス、オランダ、ベルギー、ドイツ、フランス、イタリア、ラトビア、ノルウェー、スウェーデン、)と香港、ニュージーランドだが、ロシアでも感染拡大しているのではないのか?と、推測されている。
新規感染のうち スペイン、イギリスで80% アイルランドで60%、 フランスとスイスで40% をこの変異株が占めている。
参考
「Coronavirus strain from Catalan farms accounts for large share of Europe’s new cases – study」
「Coronavirus already mutated! New virus variant is spreading in Europe」
「COVID-19ウイルスはヨーロッパで変異したか?」(英語)
③広がるミンク農場での新型コロナ感染
「20A.EU1」変異株の出発点は、スペインの北東部アルゴン州のミンク飼養農場であるとされている。
新型コロナウイルス感染が発見されたのは、スペイン東部アラゴン州テルエル県テルエル(Teruel)のミンク飼養場Secapiel社で、ミンク飼養農家の主婦に5月末感染が見つかった。
5月22日、7人が陽性となり、その後、ミンク農場でのミンクの検査が始まった。
その結果、ミンクのサンプル検査で飼養ミンクの86.67%が陽性であることがわかり、続いての同じ週に他の2人の従業員も陽性となった。
この結果を受け、92,700頭のミンクの殺処分がされた。
参考
「España sacrifica a más de 90 mil visones para prevenir el coronavirus」
新型コロナウイルスのミンクへの感染はこれが初めてではなく、オランダでは、ミンクがヒトに新型コロナウイルスを感染した疑いがあるとして、強制検査を命じ,結果、100以上のミンク農場が閉鎖されている。
オランダの2つのミンク農場では、4月23日と4月25日に最初に感染が確認された。
オランダのエラスムス大学医療センターのBas B. Oude Munninkら研究者たちは、これらのウイルスはヒトによりミンク農場に持ち込まれ、その後変異した、としている。
参考
「Scientists find evidence of two-way transmission of coronavirus on mink farms」
「Transmission of SARS-CoV-2 on mink farms between humans and mink and back to humans」
また、アメリカでもミンクへの新型コロナウイルス感染が報告されている。
ユタ、ウィスコンシン、ミシガンの各州で新型コロナでミンクが死んでおり、その総数は8月以降1万5000頭以上に及ぶ。
数十カ所の農場が調査の間隔離されている。
参考「About a dozen US mink farms in quarantine as officials probe COVID-19 outbreaks」
これらのミンクへの感染例を受け、ヨーロッパ各国やカナダ アメリカが神経を尖らせている。
カナダのミンク飼養農家は、業界への政府補償を求めているようだ。
参考「COVID-19 outbreaks at foreign mink farms have fur breeders on high alert」
これまで、ミンクに新型コロナウイルス感染がみつかったのは
デンマーク
スペイン
イタリア
オランダ
スウェーデン
アメリカ
の6カ国である。
さらにアイルランドでも、国内のミンク農場を閉鎖する必要性が叫ばれ始めている。
④デンマークの「クラスター5」変異株
ここにきて、デンマークのミンク飼養農場での新型コロナウイルス感染が顕著になってきた。
2020年11月4日、デンマーク政府は、北部ジュットランドのミンク農場にミンクを経由しての12のヒト感染があるとした。
デンマークでのコロナ感染ミンクに変異体には、5つのクラスターがある。
デンマーク州血清研究所(SSI)は、これらをクラスター1〜5として指定した。
ここには、スパイクタンパク質に次の7つの異なる変異が確認されている。
H69del / V70del、
Y453F、
I692V、
M1229I
の変異である。
参考
「All we know about mutant ‘cluster 5’ coronavirus caused by mink」
「For fear of mutated coronavirus: Denmark kills over 15 million animals」
この事態を受け、デンマーク政府は、ミンク1,700万頭を殺処分、国内で飼育されている全ミンクの殺処分を義務付けるとした命令を出したが、その後、その命令は撤回された。
感染していない農場にまでミンクの殺処分を命じる法的権限はないと判明したことを受けた措置で、改めて殺処分を義務付けるための新しい法案を議会に提出する。
何が懸念なのか?
これらの新型コロナウイルスの変異株に対して、専門家が懸念しているのは、次の点にある。
①ミンクとヒトとの相互感染
これまで、新型コロナウイルス感染は、ヒト以外の第三の中間宿主を経ずして、ヒト⇔ヒト 感染によって、拡大してきた。
であればこそ、ヒトを対象にしたワクチン開発こそが、新型コロナウイルスの感染拡大を制御しうるとされてきたのである。
ところが、これに新たに、例えば、ミンクが新型コロナウイルスの中間宿主として介在するとなった場合、これまでのヒト中心の防疫体制は、完全な見直しを迫られるはずだ。
例えば、現在、養豚の世界で蔓延している豚熱(CSF)の場合、ウイルスのホストレンジが広いために、豚にも感染するが、野生イノシシにも感染するがため、その防疫範囲は、四方八方の広さとなってしまう。
これと同じことが新型コロナウイルスにも生じてきたら、新型コロナウイルス感染による犠牲者は、膨大な数となり、ウイルス自体の根絶も、ほぼ、不可能となってしまうだろう。
それと同じ懸念を、専門家は、今回のミンクへの新型コロナウイルスの感染の広がりや、ミンクから逆にヒトが感染しているという事態に、見ているのである。
コロナウイルスは、コウモリやセンザンコウに由来するとされているが、中間宿主として哺乳類がインキュベーターとして機能し変異して、ヒトウイルスに感染しやすくなる可能性がある。
猫、犬、猿、ハムスター、フェレット、ウサギはすべて、コロナウイルスや新しいコロナウイルスからの感染に対し脆弱であり、中間宿主として機能する可能性はある。
また、中国で旧コロナウイルスがジャコウネコ属ハクビシン科のパームシベット(Paradoxurinae)によって感染拡大したことも知られている。
②ワクチン回避変異をしていないか?
デンマークの「クラスター5」と呼ばれる新型コロナウイルス変異株に対する専門家の懸念は、その変異の中に「Vaccine-escape mutations 」(ワクチン回避変異)があるかどうか?ということのようだ。
これは、先に書いたヒューストン・メソゾジストの研究者たちが懸念した「エスカピアンス」(escapians)と呼ばれるワクチンを逃れることができる突然変異体ウイルスの登場への懸念でもある。
WHOの主任科学者SoumyaSwaminathan氏は、クラスター5の突然変異が、現在開発中のワクチンの有効性にどのような影響を与える可能性があるかを言うのは時期尚早だと述べた。
しかし、専門家の懸念は、今回の「クラスター5」変異がスパイクタンパク質にあったことにある。
現在開発中のワクチンのほとんどがスパイクタンパク質の無効化に焦点を合わせているため、今回のクラスター5におけるスパイクたんぱく質での変異は現在開発中の将来のワクチンに問題を引き起こす可能性があるかもしれないとの懸念を持っているのだ。
参考「‘Mutant coronavirus’ seen before on mink farms, say scientists」
参考「The coronavirus is mutating — does it matter?」
おわりに
世界は、いよいよ、懸念されていた新型コロナウイルス蔓延の第三の波が押し寄せつつある。
この11月にロックダウンを決めた国は、イギリス、ドイツ、フランス、イタリー、スペイン、ギリシャ、ポルトガル、オーストリアの各国に及ぶ。
参考「Europe’s pandemic woes multiply with leaders drifting to lockdowns」
この上に、さらなる変異株が事態を混乱させないことを祈るばかりである。
2020/11/09記 笹山登生
各国開発の新型コロナウイルスワクチンが第3相試験を終え、次々と上市されていく雲行きだが、臨床試験不十分のワクチンが、ADE(抗体依存性感染増強)(antibody-dependent enhancement)現象を誘発する懸念が出てきた。
以下、メモ的に、順を追って詳述してみたい。
1.ADE(抗体依存性感染増強)現象とは何か?
(1)ADE(抗体依存性感染増強)とは?
ADE(抗体依存性感染増強)(antibody-dependent enhancement)とは
最初に獲得した不完全中和抗体を介し、その中和力はないがFc受容体への吸着力はあるという特性を利用(抗体依存性)して、
次に侵入した抗原(ウイルス)が、不完全中和抗体と複合体を組み、Fc受容体経路で細胞に入り込み、
細胞内で複製力を高める(感染増強)現象
をいう。
具体的には、
①新しい抗原(ウイルス)が、過去に出来ている「中和能力はないが吸着力のある不完全な中和抗体」と結びつき、
②新しい抗原と既成の不完全中和抗体との複合体が出来、
③単球またはマクロファージなどの免疫細胞を呼び寄せる不完全抗体の力を利用しながら、マクロファージの飲作用を利用して、Fc受容体を通過して細胞の中にはいりこみ、
④いったん細胞の中に入り込んだら、ウイルスの複製がうまくいくように、マクロファージのシグナル特性を、上方制御と下方制御の組み合わせで変えさせながら
ウイルス複製環境を増強していく
現象である。
このように、ADE(抗体依存性感染増強)現象は、本来は、宿主を保護するために作られたはずの抗体が、不完全な中和能力のものであったがために、再感染時において、今度は、宿主を助けるのではなく、ウイルスの複製を助ける側に回ってしまうという、きわめて、宿主にとってリスクの多い現象である。
ましてや、そのリスクの多い再感染が、本来は、宿主を救うはずのワクチン接種によっても、もたらされる危険性がある、ということである。
(2)抗体依存性感染増強(ADE)現象の歴史
A.ホークスの報告書がADE現象認識の始まり
ADE現象が認識された最初は、1964年のホークス(Hawkes RA. )による報告であった。
(参考 Hawkes RA. 「Enhancement of the infectivity of arboviruses by specific antisera produced in domestic fowls. 」Aust. J. Exp. Biol. Med. Sci.42, 465–482 (1964))
ホークスはこの報告書の中で、アルボウイルス(arbovirus)の感染力が、特定の抗血清の作用によって、見かけの感染力より増加していると指摘した。
アルボウイルスは、節足動物媒介性ウイルスともいわれ、蚊やダニなどの節足動物を介して,吸血 により脊椎動物に伝播されるウイルスである。
フラビウイ ルス科,トガウイルス科,ブニヤウイルス科,レオウイル ス科の多岐に属するウイルスである。
日本脳炎ウイルス(Japanese encephalitis virus)、西ナイルウイルス(West Nile virus)、ゲタウイルス( Getah virus)、マレーバレー脳炎ウイルス(Encephalitis Virus, Murray Valley)、などがあげられている。
B.いろいろなウイルス感染にもADE現象が
その後、1960年から1970年にかけて、同様の現象は、フラビウイ ルス科ウイルス感染を中心にして報告されてきた。
そのほか、
アルファコロナウイルス属では「猫伝染性腹膜炎ウイルス(FIPV)」
に、
ベータコロナウイルス属では「SARS-CoV-1」と「MERS」
に
同様のADE現象が見られている。
今回発生している新型コロナウイルスは、SARSコロナウイルスやMERSコロナウイルス と同じ「コロナウイルス科ベータコロナウイルス属」に分類されている。
参考「動物とヒトのコロナウイルスー2019新型コロナウイルスの流行を受けてー」
C.なぜ新型コロナウイルスにも、ADE現象が懸念されているのか?
また、新型コロナウイルスは「エンベロープを持つRNAウイルス」でもある。
「エンベロープを持つRNAウイルス」には、新型コロナウイルスが含まれるコロナウイルス科のほかに、フラビウイルス科などが含まれる。
参考「エンベローブ(ウイルス)」
フラビウイルス属は、73のウイルスがあり、そのうち、ヒトの重篤疾患に関与しているウイルスが約40ある。
フラビウイルス属は、共通の核酸ゲノム構造(正極性、約11kilobaseの一本鎖RNA)、組織(3つの構造たんぱく質、7つの非構造タンパク質)、粒子サイズ(約500オングストローム;Å)を有している。
この中で、ADE現象に関与しているフラビウイルス属ウイルスとしては、
先に記述した、1964年にホークスが報告している「アルボウイルス(arbovirus)」
や
西ナイルウイルス(West Nile virus)、
デング熱ウイルス(Dengue virus)、
ダニ媒介性脳炎ウイルス(Tick-borne encephalitis virus)、
黄熱病ウイルス(Yellow fever virus)、
ジカウイルス( Zika virus)
などがある。
そのほか
ヒト免疫不全ウイルス(HIV)(Human Immunodeficiency Virus)、
RSウイルス(RSV)(respiratory syncytial viru)、
エボラウイルス(Ebolavirus)、
インフルエンザウイルス(influenzavirus)、
口蹄疫ウイルス(FMD)(Foot-and-mouth disease virus)、
ネコ伝染性腹膜炎ウイルス(FIPV)(feline infectious peritonitis)、
ブタ繁殖・呼吸器障害症候群ウイルス(PRRSV)(Porcine reproductive and respiratory syndrome)
にも、ADE現象の発生がこれまで報告されている。
それ故に、新型コロナウイルスにも、ADE現象が懸念されているのである。
D.デング熱ウイルスからの教訓
なお、新型コロナウイルスの祖先ウイルスとみられているヒトコロナウイルスにはこれまで、下記のものが発見されている。
αコロナウイルス(γ祖先ウイルスに遠い)-HCoV-229E (1960年代に発見)、HCoV-NL63(2000年代に発見)、
βコロナウイルス(γ祖先ウイルスに近い)-SARS-CoV-2(2019年11月発見)、MERS-CoV(2012年9月発見)、SARS-CoV(2002年11月発見)、HCoV-OC43 (1960年代に発見)、HCoV-HKU1(2000年代に発見)
ADEはウイルスが抗体を利用し細胞侵入をはかり、Fc受容体との相互作用を介し細胞応答を変化させる現象であるが、このことについては、すでに2014年にスコット・B・ハルステッドがデング熱ウイルスや猫伝染性腹膜炎ウイルス(FIPV)などにつき下記論文を書いている。
「Pathogenic Exploitation of Fc Activity」
デング熱については、初回の感染時と異なる血清型のウイルスに再感染した場合に、ADE現象が発生することが報告されており、一方、ネココロナウイルスについては、初回の感染時と同じ血清型のウイルスに再感染した場合に、ADE現象が発生することが報告されている。
また、原因未解明のリーシュマニア症(leishmaniasis)(サシチョウバエの吸血により発症)にも、ADE現象が発生するといわれている。
(3)ADE(抗体依存性増強)の機序はどうなっているのか?
A.コロナウイルスが細胞内に入り込む二つの経路
コロナウイルスなど、エンベローブを持つウイルスの細胞侵入過程には次の二経路がある。
①ACE2受容体経路(エンドソーム/リソソーム経路)(endosomal pathway)
②Fc受容体経路(エンドサイトーシス経路)
このうち、ADE現象がみられるのは、②の「Fc受容体経路」である。
図1.ACE2受容体経路
図2.Fc受容体経路
上記2図は「COVID-19: Immunology and treatment options」(SusannaFelsensteinaJenny A.HerbertbPaul S.McNamarabChristian M.Hedrich)を参照
B. ADE誘発は IgG→FcγR(Fcガンマ受容体)経路で起きる
FcγR(Fcガンマ受容体)経由で細胞内侵入のウイルスへの免疫応答機序は下記のとおりである。
①過去に出来た不完全な抗体の存在
宿主内には、過去の感染や接種で産生された不完全な抗体(中和能力はないが、細胞への吸着能力がある抗体)が、既にある。
②新しい抗原と過去の不完全抗体との結合
新しく細胞内に進入しようとしている抗原(新しいウイルス粒子)は、この不完全抗体と結びつき、抗原と抗体との複合体を作る。
③免疫細胞のマクロファージのFc受容体が抗原抗体複合体を認識&結合
免疫細胞であるマクロファージの表面には、Fc受容体があり、不完全中和抗体(免疫グロブリンIgG)の複合体のFc領域を捕らえる分子が出ている。
抗原・抗体の複合体から、抗体の一部を受容体が認識し、結合する。
こうして、過去の不完全な中和抗体が橋渡しをする形で、抗原(新しいウイルス)&過去に産生の不完全中和抗体(免疫グロブリンIgG)の複合体が、マクロファージの表面上のFcγ受容体(Fcガンマ受容体)に吸着&結合する。
抗原(ウイルス)は、過去に出来た不完全抗体と結合したまま、マクロファージの中に取り込まれ、このことで、マクロファージ自体がウイルスに感染してしまう。
低親和性(結合力が弱い)のFcγ受容体(Fcガンマ受容体)は、オプソニン化(抗原に抗体や補体が結合することにより抗原が食細胞に取り込まれやすくなる現象)が促進され、活性化する。
Nikolai Eroshenko等によれば、最初の感染によって誘発された抗体を、新たに侵入の抗体をIgG抗体&Fcガンマ受容体へ、免疫細胞へのウイルスの侵入を可能にするためブリッジするので、ウイルスの向性が変化するためとしている。
既存の抗体には、感染中和の力はないが、Fcガンマ受容体を介しての抗原(ウイルス)の取り込みを誘発する力がある。
そして、既存の抗体は、マクロファージなどの免疫細胞のターゲットとしての機能を果たす。
ウイルスは、宿主の持つ過去に出来た不完全な抗体を「トロイの木馬」として、免疫細胞のターゲットとして差し出し、免疫細胞の食作用のプロセスをだまして利用し、細胞内に入り込む。
④マクロファージがエンドサイトーシス(貧食作用)により複合体を飲み込む
マクロファージはエンドサイトーシス(貧食作用)によって、これら複合体を細胞内に飲み込む。
新しいウイルスは細胞内に入り込み、細胞内のウイルス・ビリオンを増やす(外生的ADE)(B細胞・液性免疫の段階での増強)
⑤本来のマクロファージの機能は?
本来は、マクロファージは次の三つの機能を持つ。
食作用(抗原-ウイルス-を摂取)
↓
抗原提示(食作用によって取り入れた異物を分解して細胞表面に表出させる。これが抗原提示。これがシグナルとなって、ヘルパーT細胞を活性化。)
↓
活性化(マクロファージはT細胞の生産するサイトカインを受け取ることにより活性化)
⑥そして、補体がマクロファージを補助しウイルスを加水分解するはずだが?
この免疫細胞(マクロファージ)の貧食作用を補助するものとして補体があり、マクロファージが食作用で取り込んだウイルスを、補体が、加水分解していく。
本来は、抗原と複合体を形成したIgGにタンパク質複合体C1qが結合し、それをトリガーに「C1→C4→C2→C3b」の経路(古典的経路)でタンパク質分解酵素の生成で、エンべロープウイルスの周りを囲んでいる脂質層の蛋白質を加水分解していく。
なお、第三成分 (C3)が分解されるまでの補体系の活性化には、次の三つのプロセスがある。
古典経路=抗体が関与。補体第一成分 (C1) がC1qを介して結合。
副経路(第二経路)=抗体は関与しない。幾つかの成分の関与でC3が活性化。
マンノース結合レクチン(MBL)経路=MBL(またはMBP (マンナン結合蛋白))が病原体表面の糖鎖に結合して活性化。
⑦補体のエフェクター機能発動までの機序
ADE現象においては、具体的には、以下の図で
a.b.抗原・抗体の複合体にC1qが結合→c.補体のC3.C3a.C5aが活性化、膜侵襲複合体(MAC)が活性化→d.e. IgG Fc受容体は、骨髄細胞上のFcγRに結合し、エフェクター機能を発動させる。→
C3とC5aは骨髄細胞による食作用を促進→d.マクロファージ&樹状細胞による抗体依存性食作用が起きる。→e.ナチュラルキラー(NK)細胞により抗体依存性細胞傷害が媒介。→f.食細胞によるウイルス&ウイルス感染細胞の取り込み後、抗原提示、抗ウイルスT細胞活性化
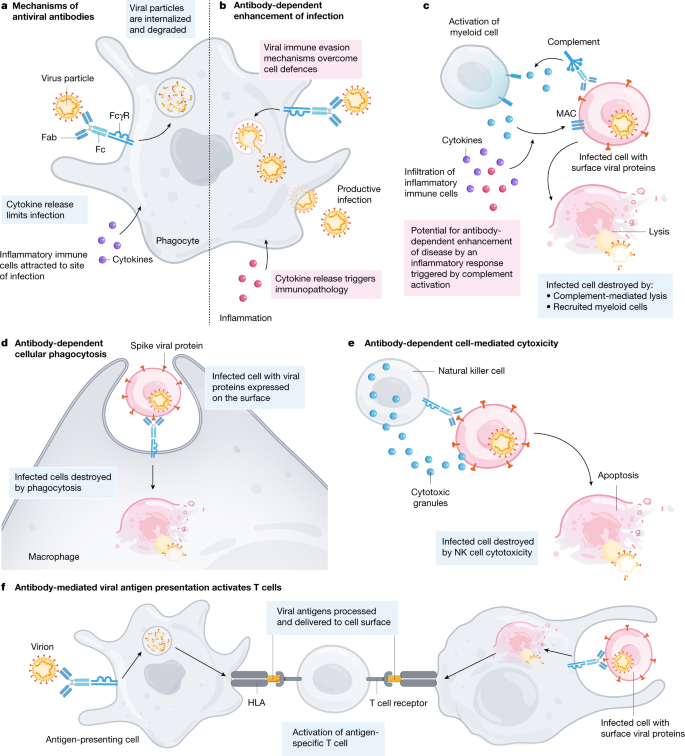
参考「A perspective on potential antibody-dependent enhancement of SARS-CoV-2」
⑧マクロファージがウイルスに感染し、本来発すべきシグナルを変質させる
ところが、ウイルスに感染したマクロファージは、本来のシグナルを変質させていく。
本来は、複合体を認識したマクロファージは、抗原提示のシグナルを出し、飲み込まれたウイルスは、補体の力を借りて、免疫細胞内で破壊&排除されるはずであるが、細胞内に入ったウイルスがマクロファージに感染(免疫細胞の感染)し、そのマクロファージの出すシグナルを変質させ、自然免疫を抑制させていく。
このマクロファージのシグナルの可逆的な変質を「マクロファージの極性化(Polarization)」という。
マクロファージのシグナル機能が可逆的な極性化によって再プログラム化され、それまでの炎症性細胞から、調節性細胞へ移行していく。
(内生的ADE)(T細胞・細胞性免疫の段階での増強)
⑨マクロファージのシグナル機能の極性化による自然免疫の抑制
マクロファージのシグナル機能の極性化によって、自然免疫は抑制され、結果、ウイルス産生増加は促進される。
インターロイキン10(IL-10)産生増加
ヘルパーT細胞のTh2細胞応答を偏らせる
⑩マクロファージの抗原提示シグナルの変質
マクロファージの出す抗原提示のシグナルにより
T細胞の中のヘルパーT細胞(リンパ球)に伝達。
T細胞活性化で、インターロイキンなどのサイトカインを生産・放出するが、マクロファージのシグナルの変質で、通常とは異なったものとなる。
すなわち
抗原提示のシグナルを送り、
T細胞のなかのヘルパーT細胞(リンパ球)に伝達、
T細胞活性化で、インターロイキンやリンフォカイン等のサイトカインを生産・放出させる。
⑪その結果は?-シグナルの変質化による免疫応答の遅延化で、ウイルス複製環境は増強へ-
シグナルのアップレギュレーションとダウンレギュレーションの相互作用で、免疫応答が遅延してくる。
ここで、もう一度、図2の「Fc受容体経路」に基づいてみてみよう。
マクロファージで取り込まれた抗原&抗体の複合体は、マクロファージの伝達シグナル特性を変えさせ、
①マクロファージのⅠ型IFNシグナル伝達(図のcとe)を阻害(ダウンレギュレーション)
②炎症性サイトカインIL-1.IL-6を発現可能(アップレギュレーション)
③TNFαの発現(図のf)増強(アップレギュレーション)
となり、
結果
Th1サイトカインIL2、TNF-α、IFN-γ減少
Th2サイトカインIL-10、IL-6、PGE-2、INF-α増加
となり、STAT転写因子経路を阻害し、
総合としては、ウイルス・ビリオンの複製環境体制を増強
④マクロファージがエンドサイトーシス(貧食作用)により複合体を飲み込む
マクロファージはエンドサイトーシス(貧食作用)によって、これら複合体を細胞内に飲み込む。
新しいウイルスは細胞内に入り込み、細胞内のウイルス・ビリオンを増やす(外生的ADE)(B細胞・液性免疫の段階での増強)
参考
抗体依存性増強のメカニズム
上記図でのABCD各段階での流れにもとずいて説明。
A.
ウイルス表面蛋白質が抗体と結合し、ウイルス・抗体複合体を形成A
この複合体が抗体のFc部分と、特定の細胞表面のFcR受容体と相互作用し、ウイルスの接着を強化。
B.
ウイルス表面蛋白質の抗体の結合により、補体C3が古典的経路で活性化。
補体Cと対応する受容体との間の相互作用により、「ウイルス-抗体-補体」複合体により、ウイルスの接着を強化。
補体C1の一部のC1q分子がセリンプロテアーゼプロ酵素であるC1rおよびC1sと共に機能。
次に、C1qが抗体複合体に結合すると、C1rおよびC1sは、血漿中のC1阻害剤の作用によりC1qから離れる。
C1が補体C2とC4を切断、C1qが離れたエフェクター補体C3と細胞上の受容体との間の組み合わせを活性化。
ウイルスは補体受容体に結合。
C.
ウイルス-抗体複合体はC1qにより結合。
C1qと受容体と結合・沈着化によりウイルスカプセルと細胞膜との融合を活性化。
D.
ウイルスの特性変化により、上方制御と下方制御の組み合わせにより、抗ウイルス遺伝子の発現が抑制。
腫瘍壊死因子(TNF)や誘導一酸化窒素シンターゼなどの抗ウイルス遺伝子の抑制により、免疫応答回避。
E.
ウイルスが抗体と結合することにより、ウイルス蛋白質のコンフォメーション(タンパク質高次構造)が変化。
ウイルスと細胞との融合が強化。
参考「Antibody-dependent enhancement of coronavirus」
C.抗原と不完全中和抗体との複合体が出来るまで
a.Fc受容体とは?
抗体のFc領域が担う機能をエフェクター機能という。
Fc領域はそれぞれに特異的なFc受容体に結合し,Fc受容体をもつ細胞を活性化したり,抗体の細胞間トランスポートに働く.
Fc受容体は、
マクロファージや単球などの食細胞、
好中球や好酸球などの顆粒球、
自然免疫系(ナチュラルキラー細胞)や適応免疫系(B細胞など)のリンパ球
など、
免疫系の多くの細胞に見られる。
Fc受容体は、抗体に結合することを可能にする。
抗体に結合し、食細胞の活性化によって、細胞が抗原を排除するのを助ける。
Fc受容体は、Bリンパ球、マクロファージ、ナチュラルキラー細胞、好中球、マスト細胞などの細胞の表面上に存在する。
b. Fc受容体に対応する抗体
Fc受容体に対応する抗体の構造は
Y字型をしており、
F(ab)「アーム」という2本の部位は、抗原への結合領域
Fc「テイル」という1本の部位は
①Fc受容体への結合領域
と
②Fc領域
からなる。
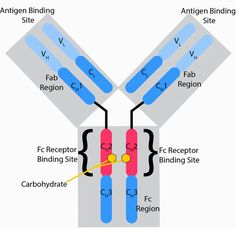
c.5つの免疫グロブリン分子に対する、それぞれ5つのFc受容体
Fc「テイル」の受容体となるFc受容体には次の5つのサブクラスがあり、
5つの免疫グロブリン分子に対しそれぞれ5つのFc受容体
が対応している。
免疫グロブリン分子→Fc受容体
①IgG→FcγR(Fcガンマ受容体)
(IgGは血中で最も豊富に認められる抗体。産生されるサイトカインの種類によって、4種類のIgGのサブクラスがある。
IgG1およびIgG3は、食細胞のFc受容体に高い親和性を示す。
食細胞のFc受容体経由オプソニン化(抗原に抗体や補体が結合することにより抗原が食細胞に取り込まれやすくなる現象)に有効。
Fc領域を介してナチュラルキラー細胞にも結合することができる。
抗体依存性の細胞毒性に寄与)
②IgE→FcεR(Fcイプシロン受容体)
(IgEは粘膜表面、血液、組織において機能)
③IgD→FcδR(Fcデルタ受容体)
(IgDはほとんどのBリンパ球の表面に存在)
④IgM→FcμR(Fcミュー受容体)
(IgMは新生児が最初に合成する免疫グロブリン)
⑤IgA→FcαR(Fcアルファ受容体)
(IgAは中和抗体としての作用)
参考「抗体の仕組みと種類を理解しよう」
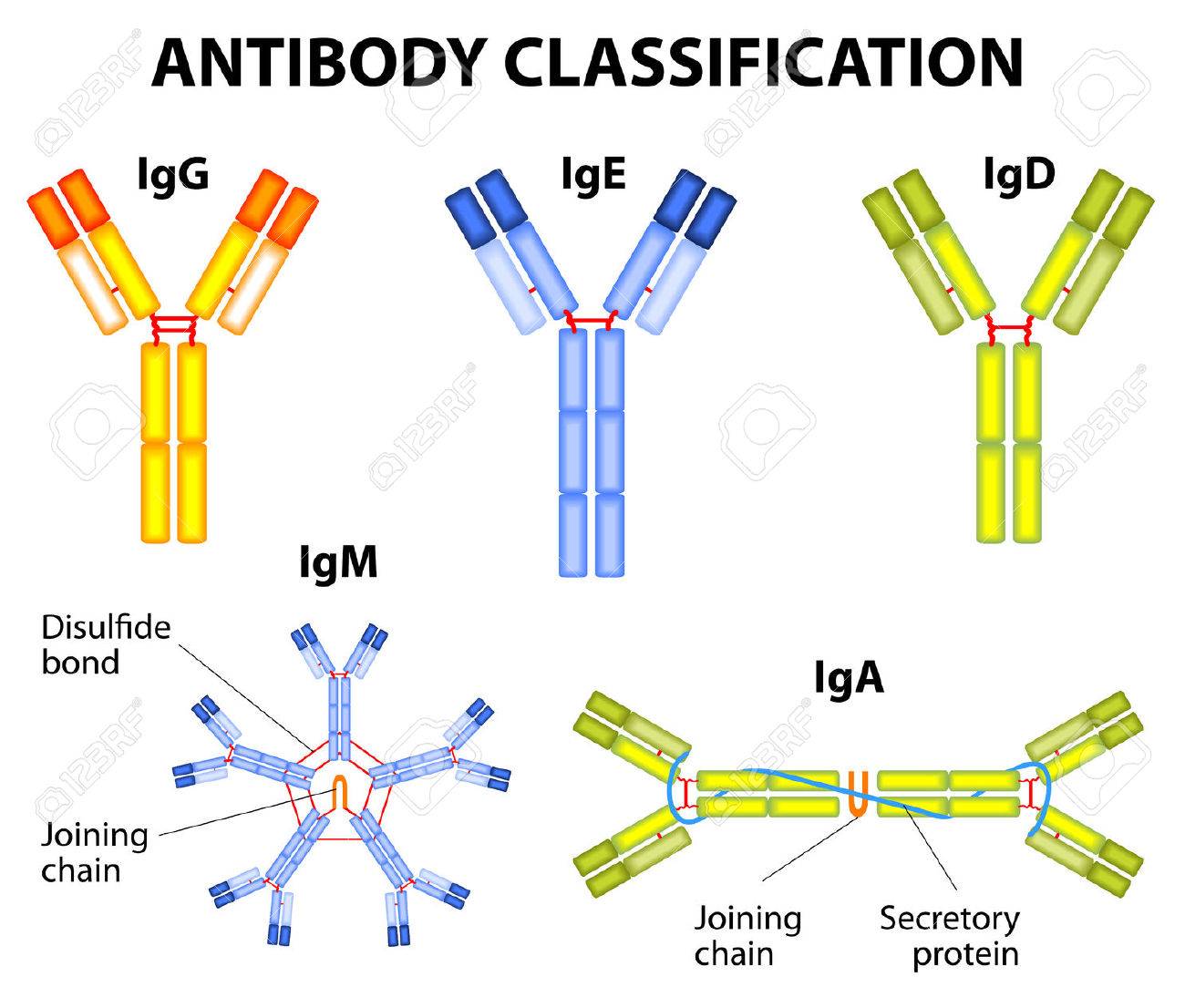
d.ADE誘発に関わるFcγR(Fcガンマ受容体)に対する免疫グロブリン分子はIgG
ADE誘発に関係のFc受容体はFcγR(Fcガンマ受容体)で免疫グロブリン分子IgGの受容体である。
なお、新生児の抗体であるIgMも、ADEを誘発しうるとの説がある。
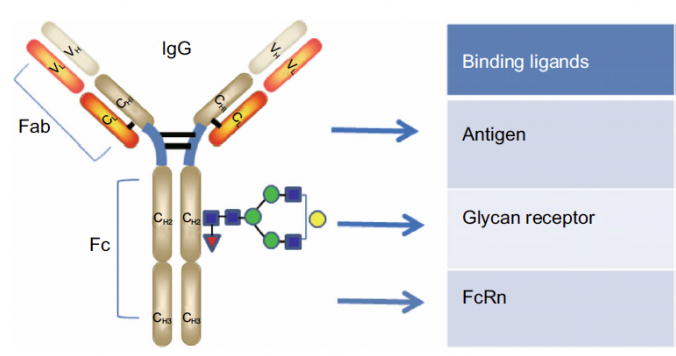
結合リガンド(特異結合物質)
は、上から
→抗原(Antigen)と結合
→糖鎖修飾誘導体(Glycan receptor)と結合
→Fc受容体(FcR)と結合
参考「Pharmacokinetics of monoclonal antibodies and Fc-fusion proteins」
免疫応答を引き出す新しいウイルス抗原は既存の抗体の免疫グロブリン分子IgGと結合し、抗原と抗体の複合体完成でIgGのY字の根元部分で受容体に結合する。
ADE現象を誘発する抗体は
F(ab)の結合力が弱いが、Fcの結合力が強いので、免疫細胞を捕らえてしまう。
これには、Fc部位の糖化状態が、関係しているとの説もある。
IgG応答は、ウイルス検出後、7日後から観察され、長期間にわたり、高いレベルで続くことがある。
なお、上記「糖鎖修飾誘導体(Glycan receptor)と結合」がADE現象に与える影響については、研究文献が少ないが、例えば、代表的な翻訳語修飾としてある「リン酸化」や「フコース」がウイルス核内移行やサイトカイン産生などにどう影響しているのか?については、興味のあるところである。
D. ACE2受容体経路は、ADEには無関係なのか?
ACE10経路の受容体は、スパイクタンパク質主体の受容体ではあるが、単球・マクロファージの受容体としては、発現が低い。
しかし、スパイクタンパク質の表面のエピトープ(抗体が認識する抗原の一部分)の異なる部分の認識によって不完全中和抗体が出来ている可能性は十分ある。
ましてや、現在世界で開発の新型コロナウイルスワクチンは、ほとんどが、ウイルスのスパイクタンパク質を標的にしたものである。
十分な力価を持ち、十分な中和抗体を形成するワクチンなら、ACE2受容体へのS蛋白質結合をブロックしうる。
本来は、スパイク蛋白質とその免疫は防御免疫応答を誘発しうるはずである。
しかし、中途半端な力価のワクチンが交差反応を起こして、不十分な中和抗体を形成し、これが、新規のウイルスの抗原&抗体複合体形成のための標的になり、FcγR(Fcガンマ受容体)経由で細胞に侵入してくる機会は十分想定しうる。
これらの複合体は、抗体力はないが、受容体への吸着力はある。
抗スパイクIgGの存在が、マクロファージ/単球におけるFcγRIIシグナル伝達を介して、免疫応答を歪めることで、ADE感染増強に関連している可能性は考えられる。
E.過炎症の指標
ADE現象の結果として起こりうるサイトカインストームによる過炎症(またはさらに超炎症;Hyper inflammation)の指標として、次のものがあげられている。
白血球(WBC)数
好中球
リンパ球数
好中球/リンパ球比(NLR)
血小板/リンパ球比(PLR)
このうち、好中球/リンパ球比(NLR)は、全身性炎症のマーカーとして、注目されている。
この比率が3を超えると、リンパ球の相対的減少状態にあるとされ、細胞性免疫の低下とみなされる・
新型コロナウイルス感染による重症化の指標とされている。
2.「抗体依存性感染増強」(ADE)現象を防ぐには?
(1)ADE誘発の最大の原因は、過去から存在していた不十分な中和抗体の存在
これまで見たように、ADE誘発の最大の原因は、次のような不完全中和抗体の存在である。
①過去の中途半端なウイルス感染や質の悪いワクチン接種によって出来た力価の不十分な中和抗体
②異種ウイルスに対する交差反応によって出来た不十分な中和抗体
③スパイクタンパク質の表面のエピトープの異なる部分の認識によって、過去に出来た不完全中和抗体
④他のコロナウイルス(先祖ウイルスかもしれないコウモリコロナウイルス)のプライミング(初回感染曝露時に免疫系を賦活するための予備刺激or準備刺激or初回刺激)によって発生した不完全中和抗体
⑤T細胞のTh2応答を刺激したかもしれない、過去に接種の麻疹ワクチンによって産生されたワクチン抗体
⑥血清型が異なるために、単に亜中和されただけの不完全中和抗体
⑦風土病化した不顕性感染のウイルスに曝露されてきた宿主が持っている不完全中和抗体
の存在である。
これらの存在が、ウイルスへの再感染やワクチンの再接種によって、ADE現象を生み出す要因になっている。。
これらは、中和抗体を作る能力はないが、Fc受容体に結合する能力がある。
そのために、ADE現象を引き起こしてしまっている。
(2).モノクローナル抗体とADEの関係は?
現在、新型コロナウイルスに対するモノクローナル抗体(mAb)としては、次の8つが研究されており、それぞれ臨床試験の段階にある。
①REGN10933+REGN10987(アメリカ)
Regeneron/N IAD
②LY3819253(LY-CoV555)(カナダ/アメリカ)
AbCellera/Eli Lilly
③JS016(中国/アメリカ)
Junshi Biosciences (上海君实生物医药科技股份有限公司)/Institute of Microbioligy.Chinese Academy of Sciences/Elilily
④TY027(シンガポール)
Tychan
⑤CT-P59(シンガポール)
Celltrion
⑥BRII-196(中国/アメリカ)
Brii Bio/TSB Therapeutics/Tsinghua University(清華大学)/The 3rd People’s Hospital of Shenzhen(深圳市第三人民医院)
⑦BRII-198(中国/アメリカ)
Brii Bio/TSB Therapeutics/Tsinghua University(清華大学)/The 3rd People’s Hospital of Shenzhen(深圳市第三人民医院)
⑧SCTA01(中国)
Sinocelltech Ltd(神州细胞)/Chinese Academy of Sciences(中国科学院)
参考
「Clinical studies evaluating anti-SARS-CoV-2 monoclonal antibodies」
これらのmAbの投与が、ADEの原因につながる不完全抗体を生みうるかどうか?については、今後の研究が必要のようである。
ちなみに、デング熱については、哺乳類細胞で産生されたmAb(mammalian-cell-produced)は、ADE誘発の懸念があるが、植物細胞で産生されたmAb(plant-based)は、ADE誘発の危険性は少ない、との研究結果はすでにある。
(3)ADE誘発は細胞性免疫の誘導失敗が原因
では、どうやってADE誘発リスクをコントロールするのか?
免疫は、そのメカニズムによって2つに大別される。
1つは、液性免疫と呼ばれ、ワクチンの接種や過去の感染によって体内で作られる抗体が活躍する。
抗体には、病原体などに結合することで感染力や毒性を失わせる作用を持つものや、マクロファージなどの免疫細胞による取り込み・処理を助けるものなどがある。
2つは、細胞性免疫と呼ばれ、マクロファージやキラーT細胞といった免疫細胞が病原体や病原体に感染した細胞を直接取り込み(貪食と呼ばれる)、処理することで体を守るシステムである。
細胞性免疫はマクロファージ、キラーT細胞等免疫細胞が感染細胞を貪食で守る。
ADE誘発は、新しいウイルスが、液性免疫の機序を利用して、過去に出来ていた「中和能力はないが吸着能力のある抗体」と複合体を形成し、細胞の中に入り込んだ。
過去の液性免疫の存在が、ADE現象を生み出すもともとの原因であるともいえる。
その次に、免疫細胞であるマクロファージの可逆的極性化特性を利用して、マクロファージのシグナル特性を、ダウンレギュレートの方向に替え、ウイルスの複製環境を整えた。
この後者のマクロファージの特性を変えさせたのは、細胞性免疫の誘導に失敗したためであり、そこにADE現象は起こっている。
ヘルパーT細胞のうち
細胞性免疫の調節がTh1
抗体産生など液性免疫の調節がTh2
で行われるが、ADE現象は、Th1のアップレギュレーションによって、腫瘍壊死因子であるTNF-αの増強を許してしまっていることによる。
ADE現象の発生を回避するには、
細胞免疫の調整を司るTh1と
液性免疫の調整を司るTH2と
のバランスが重要になってくる。
これらの宿主にとっての好ましくない免疫応答が「外生(液性免疫)と内生(細胞性免疫)との二段階」で増強される現象が生じてくる。
図 内生(細胞性免疫)ADE
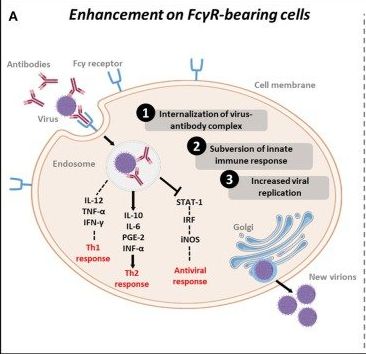
機序
①抗原と抗体との複合体は、免疫細胞上のFc領域へ結合後、細胞内に内在化
②Th1サイトカインのIL2、TNF-α、IFN-γ を減少させる。
Th2サイトカインのIL-10、IL-6、PGE-2、INF-α を増加させる。
結果、STAT経路を阻害し、免疫応答レベルを低下させる。
③抗ウイルス応答の抑制により、結果として、ウイルス複製は増加。
図 外生(液性免疫)ADE
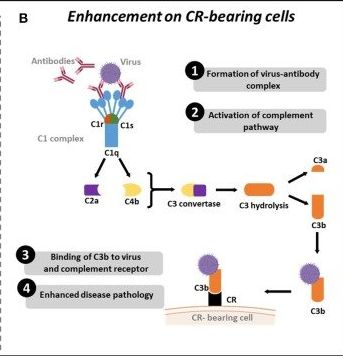
機序
①抗原&抗体の複合体形成
②複合体が免疫複合体と結合し、補体経路を活性化
③補体フラグメントC4bとC2aとが動員され、C3転換酵素を生成、次にC3が加水分解され、C3bが生成
④C3bは、ウイルスに結合し、CR保有細胞の受容体(CR)を補う
⑤細胞の溶解をし、疾患の悪化を促進させる。
上記図は「Viral-Induced Enhanced Disease Illness」より引用
3.ADEを誘発させない新型コロナウイルスワクチンの条件とは?-終わりに-
ここで、ワクチン接種によって十分な中和抗体が出来るか?が問題となる。
ADE応答は抗体価に依存する。
抗体が低濃度で存在の場合はADEが発生するが、高濃度で存在の場合にADEが発生しない。
また、中和抗体の親和性(結合力)が低い場合にも、ADE現象は発生する。
理 想 的 なワクチンは、SARS-CoV-2 に対して特異的かつ効率的に作用する中和抗体を産生するものでなければならない。
しかし、一定のワクチンの中には、中和抗体の力価が不十分であるために、抗体依存性増強(ADE)が引き起こされる可能性があるワクチンもありうる。
また、ADEと似た症状に「ワクチン過敏反応」(VAH)があるが、これとADE現象と混同しない対応が求められる。
ワクチン抗体産生が不十分で産生された抗体は、新しい感染ウイルスとが結合し、免疫複合体を生成し、Fc受容体経由で、細胞の中に進入する。
十分な力価のないワクチンで産生された抗体、または再感染ウイルスと免疫複合体を作り、ACE2受容体経由でなく、FcγR(Fcガンマ受容体)経由で、マクロファージによる貪食作用で細胞内に進入する。
これらのことから言えば、理想的な新型コロナウイルスワクチンは、「抗体が関与する液性免疫を誘導することなく、細胞性免疫を優位に誘導するワクチン」ということになる。
ワクチン設計にあたり、中和抗体と強力なT細胞性免疫の両方を誘導でき、かつ、ADE誘導性、非中和性、または弱中和性のエピトープを除外することが必要になりうる。
また、中和抗体の出来にくい低力価のワクチンはADE誘発を招きかねず、そのためには、最初のワクチン接種で高力価を保証するための、アジュバントの開発も必要となりうる。
ある識者は、
「S蛋白質に対する不完全な免疫(抗体)が誘導されれば、ADEが起こる可能性がある。
細胞性免疫を誘導するワクチンの開発は、(ADEを防ぐための)1つの手段になり得る。
ウイルスのS蛋白質のうち、感染において特に重要な役割を担う一部の領域をターゲットにしたワクチンを開発することで、不要なエピトープの認識することなく、そのことで、不要な不完全S抗体を生むこともなくなる」
と指摘している。
新型コロナウイルスの不完全抗体にもADE現象誘発の懸念があるかどうかは、まだ、わかっていないが、つい最近、中国の復旦大学のHuang氏らが、次のような報告をしている。
特定の新型コロナウイルス患者に「7F3」と呼ばれる高リスク抗体(ADE現象を誘発)を発見したとして、222人の患者のうち、軽度症状者の8%、回復者の76%に「7F3」抗体が見つかったとして、この高リスク抗体がADE現象を誘発するのではないか?との仮説を示している。
参考
以上
参考
1.現在開発中の新型コロナウイルスワクチン一覧
2.現在開発中のモノクローナル抗体一覧
参考文献
「Antibody-Dependent Enhancement (ADE) Phenomenon Could Be Caused By SARS-COV-2 Antibody」(2020/09/29)
「Immunoglobulin G2 Antibody as a Potential Target for COVID-19 Vaccine」(2020/09/29)
「Immune Response to COVID-19: Can We Benefit from the SARS-CoV and MERS-CoV Pandemic Experience?」 (2020/09/18)
「Another study by researchers from Nanjing University Medical School 」 (2020/09/11)
「Antibody-dependent enhancement of coronavirus」(2020/09/10)
「Antibody-dependent enhancement and SARS-CoV-2 vaccines and therapies」 (2020/09/09)
「Antibody-dependent enhancement of Coronavirus」(2020/09/07 )
「Immunological considerations for COVID-19 vaccine strategies」 (2020/09/04)
「Four scenarios on how we might develop immunity to Covid-19」 (2020/08/26)
「Structural and dynamical analysis of integrated human/SARS-CoV-2 metabolic models present novel treatment strategies against COVID-19」 (2020/08/14)
「Deciphering SARS-CoV-2 Virologic and Immunologic Features」 (2020/08/14)
「COVID 19 Vaccines: Should we fear ADE? 」 (2020/08/12)
「The role of IgG Fc receptors in antibody-dependent enhancement」(2020/08/11)
「Presence of Genetic Variants Among Young Men With Severe COVID-19」 (2020/07/24)
「Immunity 101 for COVID-19」 (2020/07/18)
「A perspective on potential antibody-dependent enhancement of SARS-CoV-2」 (2020/07/13)
「COVID-19 Coronavirus Vaccine Design Using Reverse Vaccinology and Machine Learning」 (2020/07)
「Implications of antibody-dependent enhancement of infection for SARS-CoV-2 countermeasures」 (2020/06/05)
「Impact of immune enhancement on Covid-19 polyclonal hyperimmune globulin therapy and vaccine development」 (2020/05/01)
「What immunity to Covid-19 might actually mean」(2020/04/23)
「The potential danger of suboptimal antibody responses in COVID-19」(2020/04/21)
「Coronavirus could attack immune system like HIV by targeting protective cells, warn scientists」(2020/04/18)
「SARS-CoV-2 and COVID-19 in older adults: what we may expect regarding pathogenesis, immune responses, and outcomes」 (2020/04/10)
「RETRACTED ARTICLE: SARS-CoV-2 infects T lymphocytes through its spike protein-mediated membrane fusion」 (2020/04/07)
「COVID-19: Immunology and treatment options」 (2020/04)
「Hypothesis for potential pathogenesis of SARS-CoV-2 infection–a review of immune changes in patients with viral pneumonia」 (2020/03/18)
「Immune responses in COVID-19 and potential vaccines: Lessons learned from SARS and MERS epidemic 」 (2020/03)
「Coronavirus infections and immune responses」 (2020/01/25)
「Viral-Induced Enhanced Disease Illness」(2018/12/05)
「Molecular Mechanism for Antibody-Dependent Enhancement of Coronavirus Entry」(2019/12/04)
「FcγRs Muscle Up Dendritic Cell Cross-Presentation」 (2013/01/09)
「Viral Innate Immune Evasion and the Pathogenesis of Emerging RNA Virus Infections」 (2019/10/18)
「Molecular Mechanism for Antibody-Dependent Enhancement of Coronavirus Entry」 (2015)
「Antibody Dependent Enhancement」 (2014)
「Antibody identification」 (?)
「SARS-CoV-2 (COVID-19) S1 protein (D614G), His Tag」 (?)
「Immunology Research Scope」 (?)
「免疫グロブリンFc受 容体を介 した免疫抑制機構」(2000)
「フローサイトメトリー (FCM) / FACS 抗体は、Fc受容体ブロッキングによって検証されます」(?)
以上
各国開発の新型コロナウイルスワクチンが第3相試験を終え、次々と上市されていく雲行きだが、臨床試験不十分のワクチンが、ADE(抗体依存性感染増強)(antibody-dependent enhancement)現象を誘発する懸念が出てきた。
以下、メモ的に、順を追って詳述してみたい。
1.ADE(抗体依存性感染増強)現象とは何か?
(1)ADE(抗体依存性感染増強)とは?
ADE(抗体依存性感染増強)(antibody-dependent enhancement)とは
最初に獲得した不完全中和抗体を介し、その中和力はないがFc受容体への吸着力はあるという特性を利用(抗体依存性)して、
次に侵入した抗原(ウイルス)が、不完全中和抗体と複合体を組み、Fc受容体経路で細胞に入り込み、
細胞内で複製力を高める(感染増強)現象
をいう。
具体的には、
①新しい抗原(ウイルス)が、過去に出来ている「中和能力はないが吸着力のある不完全な中和抗体」と結びつき、
②新しい抗原と既成の不完全中和抗体との複合体が出来、
③単球またはマクロファージなどの免疫細胞を呼び寄せる不完全抗体の力を利用しながら、マクロファージの飲作用を利用して、Fc受容体を通過して細胞の中にはいりこみ、
④いったん細胞の中に入り込んだら、ウイルスの複製がうまくいくように、マクロファージのシグナル特性を、上方制御と下方制御の組み合わせで変えさせながら
ウイルス複製環境を増強していく
現象である。
このように、ADE(抗体依存性感染増強)現象は、本来は、宿主を保護するために作られたはずの抗体が、不完全な中和能力のものであったがために、再感染時において、今度は、宿主を助けるのではなく、ウイルスの複製を助ける側に回ってしまうという、きわめて、宿主にとってリスクの多い現象である。
ましてや、そのリスクの多い再感染が、本来は、宿主を救うはずのワクチン接種によっても、もたらされる危険性がある、ということである。
(2)抗体依存性感染増強(ADE)現象の歴史
ADE現象が認識された最初は、1964年のホークス(Hawkes RA. )による報告であった。
(参考 Hawkes RA. 「Enhancement of the infectivity of arboviruses by specific antisera produced in domestic fowls. 」Aust. J. Exp. Biol. Med. Sci.42, 465–482 (1964))
ホークスはこの報告書の中で、アルボウイルス(arbovirus)の感染力が、特定の抗血清の作用によって、見かけの感染力より増加していると指摘した。
アルボウイルスは、節足動物媒介性ウイルスともいわれ、蚊やダニなどの節足動物を介して,吸血 により脊椎動物に伝播されるウイルスである。
フラビウイ ルス科,トガウイルス科,ブニヤウイルス科,レオウイル ス科の多岐に属するウイルスである。
日本脳炎ウイルス(Japanese encephalitis virus)、西ナイルウイルス(West Nile virus)、ゲタウイルス( Getah virus)、マレーバレー脳炎ウイルス(Encephalitis Virus, Murray Valley)、などがあげられている。
その後、1960年から1970年にかけて、同様の現象は、フラビウイ ルス科ウイルス感染を中心にして報告されてきた。
そのほか、
アルファコロナウイルス属では「猫伝染性腹膜炎ウイルス(FIPV)」
に、
ベータコロナウイルス属では「SARS-CoV-1」と「MERS」
に
同様のADE現象が見られている。
今回発生している新型コロナウイルスは、SARSコロナウイルスやMERSコロナウイルス と同じ「コロナウイルス科ベータコロナウイルス属」に分類されている。
参考「動物とヒトのコロナウイルスー2019新型コロナウイルスの流行を受けてー」
また、新型コロナウイルスは「エンベロープを持つRNAウイルス」でもある。
「エンベロープを持つRNAウイルス」には、新型コロナウイルスが含まれるコロナウイルス科のほかに、フラビウイルス科などが含まれる。
参考「エンベローブ(ウイルス)」
フラビウイルス属は、73のウイルスがあり、そのうち、ヒトの重篤疾患に関与しているウイルスが約40ある。
フラビウイルス属は、共通の核酸ゲノム構造(正極性、約11kilobaseの一本鎖RNA)、組織(3つの構造たんぱく質、7つの非構造タンパク質)、粒子サイズ(約500オングストローム;Å)を有している。
この中で、ADE現象に関与しているフラビウイルス属ウイルスとしては、
先に記述した、1964年にホークスが報告している「アルボウイルス(arbovirus)」
や
西ナイルウイルス(West Nile virus)、
デング熱ウイルス(Dengue virus)、
ダニ媒介性脳炎ウイルス(Tick-borne encephalitis virus)、
黄熱病ウイルス(Yellow fever virus)、
ジカウイルス( Zika virus)
などがある。
そのほか
ヒト免疫不全ウイルス(HIV)(Human Immunodeficiency Virus)、
RSウイルス(RSV)(respiratory syncytial viru)、
エボラウイルス(Ebolavirus)、
インフルエンザウイルス(influenzavirus)、
口蹄疫ウイルス(FMD)(Foot-and-mouth disease virus)、
ネコ伝染性腹膜炎ウイルス(FIPV)(feline infectious peritonitis)、
ブタ繁殖・呼吸器障害症候群ウイルス(PRRSV)(Porcine reproductive and respiratory syndrome)
にも、ADE現象の発生がこれまで報告されている。
それ故に、新型コロナウイルスにも、ADE現象が懸念されているのである。
なお、新型コロナウイルスの祖先ウイルスとみられているヒトコロナウイルスにはこれまで、下記のものが発見されている。
αコロナウイルス(γ祖先ウイルスに遠い)-HCoV-229E (1960年代に発見)、HCoV-NL63(2000年代に発見)、
βコロナウイルス(γ祖先ウイルスに近い)-SARS-CoV-2(2019年11月発見)、MERS-CoV(2012年9月発見)、SARS-CoV(2002年11月発見)、HCoV-OC43 (1960年代に発見)、HCoV-HKU1(2000年代に発見)
ADEはウイルスが抗体を利用し細胞侵入をはかり、Fc受容体との相互作用を介し細胞応答を変化させる現象であるが、このことについては、すでに2014年にスコット・B・ハルステッドが他ウイルスにつき下記論文を書いている。
「Pathogenic Exploitation of Fc Activity」
デング熱については、初回の感染時と異なる血清型のウイルスに再感染した場合に、ADE現象が発生することが報告されており、一方、ネココロナウイルスについては、初回の感染時と同じ血清型のウイルスに再感染した場合に、ADE現象が発生することが報告されている。
また、原因未解明のリーシュマニア症(leishmaniasis)(サシチョウバエの吸血により発症)にも、ADE現象が発生するといわれている。
(3)ADE(抗体依存性増強)の機序はどうなっているのか?
A.コロナウイルスが細胞内に入り込む二つの経路
コロナウイルスなど、エンベローブを持つウイルスの細胞侵入過程には次の二経路がある。
①ACE2受容体経路(エンドソーム/リソソーム経路)(endosomal pathway)
②Fc受容体経路(エンドサイトーシス経路)
このうち、ADE現象がみられるのは、②の「Fc受容体経路」である。
図1.ACE2受容体経路
図2.Fc受容体経路
上記2図は「COVID-19: Immunology and treatment options」(SusannaFelsensteinaJenny A.HerbertbPaul S.McNamarabChristian M.Hedrich)を参照
B. ADE誘発は IgG→FcγR(Fcガンマ受容体)経路で起きる
FcγR(Fcガンマ受容体)経由で細胞内侵入のウイルスへの免疫応答機序は下記のとおりである。
①過去に出来た不完全な抗体の存在
宿主内には、過去の感染や接種で産生された不完全な抗体(中和能力はないが、細胞への吸着能力がある抗体)が、既にある。
②新しい抗原と過去の不完全抗体との結合
新しく細胞内に進入しようとしている抗原(新しいウイルス粒子)は、この不完全抗体と結びつき、抗原と抗体との複合体を作る。
③免疫細胞のマクロファージのFc受容体が抗原抗体複合体を認識&結合
免疫細胞であるマクロファージの表面には、Fc受容体があり、不完全中和抗体(免疫グロブリンIgG)の複合体のFc領域を捕らえる分子が出ている。
抗原・抗体の複合体から、抗体の一部を受容体が認識し、結合する。
こうして、過去の不完全な中和抗体が橋渡しをする形で、抗原(新しいウイルス)&過去に産生の不完全中和抗体(免疫グロブリンIgG)の複合体が、マクロファージの表面上のFcγ受容体(Fcガンマ受容体)に吸着&結合する。
抗原(ウイルス)は、過去に出来た不完全抗体と結合したまま、マクロファージの中に取り込まれ、このことで、マクロファージ自体がウイルスに感染してしまう。
低親和性(結合力が弱い)のFcγ受容体(Fcガンマ受容体)は、オプソニン化(抗原に抗体や補体が結合することにより抗原が食細胞に取り込まれやすくなる現象)が促進され、活性化する。
Nikolai Eroshenko等によれば、最初の感染によって誘発された抗体を、新たに侵入の抗体をIgG抗体&Fcガンマ受容体へ、免疫細胞へのウイルスの侵入を可能にするためブリッジするので、ウイルスの向性が変化するためとしている。
既存の抗体には、感染中和の力はないが、Fcガンマ受容体を介しての抗原(ウイルス)の取り込みを誘発する力がある。
そして、既存の抗体は、マクロファージなどの免疫細胞のターゲットとしての機能を果たす。
ウイルスは、宿主の持つ過去に出来た不完全な抗体を「トロイの木馬」として、免疫細胞のターゲットとして差し出し、免疫細胞の食作用のプロセスをだまして利用し、細胞内に入り込む。
④マクロファージがエンドサイトーシス(貧食作用)により複合体を飲み込む
マクロファージはエンドサイトーシス(貧食作用)によって、これら複合体を細胞内に飲み込む。
新しいウイルスは細胞内に入り込み、細胞内のウイルス・ビリオンを増やす(外生的ADE)(B細胞・液性免疫の段階での増強)
⑤本来のマクロファージの機能は?
本来は、マクロファージは次の三つの機能を持つ。
食作用(抗原-ウイルス-を摂取)
↓
抗原提示(食作用によって取り入れた異物を分解して細胞表面に表出させる。これが抗原提示。これがシグナルとなって、ヘルパーT細胞を活性化。)
↓
活性化(マクロファージはT細胞の生産するサイトカインを受け取ることにより活性化)
⑥そして、補体がマクロファージを補助しウイルスを加水分解するはずだが?
この免疫細胞(マクロファージ)の貧食作用を補助するものとして補体があり、マクロファージが食作用で取り込んだウイルスを、補体が、加水分解していく。
本来は、抗原と複合体を形成したIgGにタンパク質複合体C1qが結合し、それをトリガーに「C1→C4→C2→C3b」の経路(古典的経路)でタンパク質分解酵素の生成で、エンべロープウイルスの周りを囲んでいる脂質層の蛋白質を加水分解していく。
なお、第三成分 (C3)が分解されるまでの補体系の活性化には、次の三つのプロセスがある。
古典経路=抗体が関与。補体第一成分 (C1) がC1qを介して結合。
副経路(第二経路)=抗体は関与しない。幾つかの成分の関与でC3が活性化。
マンノース結合レクチン(MBL)経路=MBL(またはMBP (マンナン結合蛋白))が病原体表面の糖鎖に結合して活性化。
⑦補体のエフェクター機能発動までの機序
ADE現象においては、具体的には、以下の図で
a.b.抗原・抗体の複合体にC1qが結合→c.補体のC3.C3a.C5aが活性化、膜侵襲複合体(MAC)が活性化→d.e. IgG Fc受容体は、骨髄細胞上のFcγRに結合し、エフェクター機能を発動させる。→
C3とC5aは骨髄細胞による食作用を促進→d.マクロファージ&樹状細胞による抗体依存性食作用が起きる。→e.ナチュラルキラー(NK)細胞により抗体依存性細胞傷害が媒介。→f.食細胞によるウイルス&ウイルス感染細胞の取り込み後、抗原提示、抗ウイルスT細胞活性化
参考「A perspective on potential antibody-dependent enhancement of SARS-CoV-2」
⑦マクロファージがウイルスに感染し、本来発すべきシグナルを変質させる
ところが、ウイルスに感染したマクロファージは、本来のシグナルを変質させていく。
本来は、複合体を認識したマクロファージは、抗原提示のシグナルを出し、飲み込まれたウイルスは、補体の力を借りて、免疫細胞内で破壊&排除されるはずであるが、細胞内に入ったウイルスがマクロファージに感染(免疫細胞の感染)し、そのマクロファージの出すシグナルを変質させ、自然免疫を抑制させていく。
このマクロファージのシグナルの可逆的な変質を「マクロファージの極性化(Polarization)」という。
マクロファージのシグナル機能が可逆的な極性化によって再プログラム化され、それまでの炎症性細胞から、調節性細胞へ移行していく。
(内生的ADE)(T細胞・細胞性免疫の段階での増強)
⑧マクロファージのシグナル機能の極性化による自然免疫の抑制
マクロファージのシグナル機能の極性化によって、自然免疫は抑制され、結果、ウイルス産生増加は促進される。
インターロイキン10(IL-10)産生増加
ヘルパーT細胞のTh2細胞応答を偏らせる
⑨マクロファージの抗原提示シグナルの変質
マクロファージの出す抗原提示のシグナルにより
T細胞の中のヘルパーT細胞(リンパ球)に伝達。
T細胞活性化で、インターロイキンなどのサイトカインを生産・放出するが、マクロファージのシグナルの変質で、通常とは異なったものとなる。
すなわち
抗原提示のシグナルを送り、
T細胞のなかのヘルパーT細胞(リンパ球)に伝達、
T細胞活性化で、インターロイキンやリンフォカイン等のサイトカインを生産・放出させる。
⑩その結果は?-シグナルの変質化による免疫応答の遅延化で、ウイルス複製環境は増強へ-
シグナルのアップレギュレーションとダウンレギュレーションの相互作用で、免疫応答が遅延してくる。
ここで、もう一度、図2の「Fc受容体経路」に基づいてみてみよう。
マクロファージで取り込まれた抗原&抗体の複合体は、マクロファージの伝達シグナル特性を変えさせ、
①マクロファージのⅠ型IFNシグナル伝達(図のcとe)を阻害(ダウンレギュレーション)
②炎症性サイトカインIL-1.IL-6を発現可能(アップレギュレーション)
③TNFαの発現(図のf)増強(アップレギュレーション)
となり、
結果
Th1サイトカインIL2、TNF-α、IFN-γ減少
Th2サイトカインIL-10、IL-6、PGE-2、INF-α増加
となり、STAT転写因子経路を阻害し、
総合としては、ウイルス・ビリオンの複製環境体制を増強
C.抗原と不完全中和抗体との複合体が出来るまで
a.Fc受容体とは?
抗体のFc領域が担う機能をエフェクター機能という。
Fc領域はそれぞれに特異的なFc受容体に結合し,Fc受容体をもつ細胞を活性化したり,抗体の細胞間トランスポートに働く.
Fc受容体は、
マクロファージや単球などの食細胞、
好中球や好酸球などの顆粒球、
自然免疫系(ナチュラルキラー細胞)や適応免疫系(B細胞など)のリンパ球
など、
免疫系の多くの細胞に見られる。
Fc受容体は、抗体に結合することを可能にする。
抗体に結合し、食細胞の活性化によって、細胞が抗原を排除するのを助ける。
Fc受容体は、Bリンパ球、マクロファージ、ナチュラルキラー細胞、好中球、マスト細胞などの細胞の表面上に存在する。
b. Fc受容体に対応する抗体
Fc受容体に対応する抗体の構造は
Y字型をしており、
F(ab)「アーム」という2本の部位は、抗原への結合領域
Fc「テイル」という1本の部位は
①Fc受容体への結合領域
と
②Fc領域
からなる。
c.5つの免疫グロブリン分子に対する、それぞれ5つのFc受容体
Fc「テイル」の受容体となるFc受容体には次の5つのサブクラスがあり、
5つの免疫グロブリン分子に対しそれぞれ5つのFc受容体
が対応している。
免疫グロブリン分子→Fc受容体
①IgG→FcγR(Fcガンマ受容体)
(IgGは血中で最も豊富に認められる抗体。産生されるサイトカインの種類によって、4種類のIgGのサブクラスがある。
IgG1およびIgG3は、食細胞のFc受容体に高い親和性を示す。
食細胞のFc受容体経由オプソニン化(抗原に抗体や補体が結合することにより抗原が食細胞に取り込まれやすくなる現象)に有効。
Fc領域を介してナチュラルキラー細胞にも結合することができる。
抗体依存性の細胞毒性に寄与)
②IgE→FcεR(Fcイプシロン受容体)
(IgEは粘膜表面、血液、組織において機能)
③IgD→FcδR(Fcデルタ受容体)
(IgDはほとんどのBリンパ球の表面に存在)
④IgM→FcμR(Fcミュー受容体)
(IgMは新生児が最初に合成する免疫グロブリン)
⑤IgA→FcαR(Fcアルファ受容体)
(IgAは中和抗体としての作用)
参考「抗体の仕組みと種類を理解しよう」
d.ADE誘発に関わるFcγR(Fcガンマ受容体)に対する免疫グロブリン分子はIgG
ADE誘発に関係のFc受容体はFcγR(Fcガンマ受容体)で免疫グロブリン分子IgGの受容体である。
なお、新生児の抗体であるIgMも、ADEを誘発しうるとの説がある。
結合リガンド(特異結合物質)
は、上から
→抗原(Antigen)と結合
→糖鎖修飾誘導体(Glycan receptor)と結合
→Fc受容体(FcR)と結合
参考「Pharmacokinetics of monoclonal antibodies and Fc-fusion proteins」
免疫応答を引き出す新しいウイルス抗原は既存の抗体の免疫グロブリン分子IgGと結合し、抗原と抗体の複合体完成でIgGのY字の根元部分で受容体に結合する。
ADE現象を誘発する抗体は
F(ab)の結合力が弱いが、Fcの結合力が強いので、免疫細胞を捕らえてしまう。
これには、Fc部位の糖化状態が、関係しているとの説もある。
IgG応答は、ウイルス検出後、7日後から観察され、長期間にわたり、高いレベルで続くことがある。
なお、上記「糖鎖修飾誘導体(Glycan receptor)と結合」がADE現象に与える影響については、研究文献が少ないが、例えば、代表的な翻訳語修飾としてある「リン酸化」や「フコース」がウイルス核内移行やサイトカイン産生などにどう影響しているのか?については、興味のあるところである。
D. ACE2受容体経路は、ADEには無関係なのか?
ACE10経路の受容体は、スパイクタンパク質主体の受容体ではあるが、単球・マクロファージの受容体としては、発現が低い。
しかし、スパイクタンパク質の表面のエピトープ(抗体が認識する抗原の一部分)の異なる部分の認識によって不完全中和抗体が出来ている可能性は十分ある。
ましてや、現在世界で開発の新型コロナウイルスワクチンは、ほとんどが、ウイルスのスパイクタンパク質を標的にしたものである。
十分な力価を持ち、十分な中和抗体を形成するワクチンなら、ACE2受容体へのS蛋白質結合をブロックしうる。
本来は、スパイク蛋白質とその免疫は防御免疫応答を誘発しうるはずである。
しかし、中途半端な力価のワクチンが交差反応を起こして、不十分な中和抗体を形成し、これが、新規のウイルスの抗原&抗体複合体形成のための標的になり、FcγR(Fcガンマ受容体)経由で細胞に侵入してくる機会は十分想定しうる。
これらの複合体は、抗体力はないが、受容体への吸着力はある。
抗スパイクIgGの存在が、マクロファージ/単球におけるFcγRIIシグナル伝達を介して、免疫応答を歪めることで、ADE感染増強に関連している可能性は考えられる。
E.過炎症の指標
ADE現象の結果として起こりうるサイトカインストームによる過炎症(またはさらに超炎症;Hyper inflammation)の指標として、次のものがあげられている。
白血球(WBC)数
好中球
リンパ球数
好中球/リンパ球比(NLR)
血小板/リンパ球比(PLR)
このうち、好中球/リンパ球比(NLR)は、全身性炎症のマーカーとして、注目されている。
この比率が3を超えると、リンパ球の相対的減少状態にあるとされ、細胞性免疫の低下とみなされる・
新型コロナウイルス感染による重症化の指標とされている。
2.「抗体依存性感染増強」(ADE)現象を防ぐには?
(1)ADE誘発の最大の原因は、過去から存在していた不十分な中和抗体の存在
これまで見たように、ADE誘発の最大の原因は、次のような不完全中和抗体の存在である。
①過去の中途半端なウイルス感染や質の悪いワクチン接種によって出来た力価の不十分な中和抗体
②異種ウイルスに対する交差反応によって出来た不十分な中和抗体
③スパイクタンパク質の表面のエピトープの異なる部分の認識によって、過去に出来た不完全中和抗体
④他のコロナウイルス(先祖ウイルスかもしれないコウモリコロナウイルス)のプライミング(初回感染曝露時に免疫系を賦活するための予備刺激or準備刺激or初回刺激)によって発生した不完全中和抗体
⑤T細胞のTh2応答を刺激したかもしれない、過去に接種の麻疹ワクチンによって産生されたワクチン抗体
⑥血清型が異なるために、単に亜中和されただけの不完全中和抗体
⑦風土病化した不顕性感染のウイルスに曝露されてきた宿主が持っている不完全中和抗体
の存在である。
これらの存在が、ウイルスへの再感染やワクチンの再接種によって、ADE現象を生み出す要因になっている。。
これらは、中和抗体を作る能力はないが、Fc受容体に結合する能力がある。
そのために、ADE現象を引き起こしてしまっている。
(2).モノクローナル抗体とADEの関係は?
現在、新型コロナウイルスに対するモノクローナル抗体(mAb)としては、次の8つが研究されており、それぞれ臨床試験の段階にある。
①REGN10933+REGN10987(アメリカ)
Regeneron/N IAD
②LY3819253(LY-CoV555)(カナダ/アメリカ)
AbCellera/Eli Lilly
③JS016(中国/アメリカ)
Junshi Biosciences (上海君实生物医药科技股份有限公司)/Institute of Microbioligy.Chinese Academy of Sciences/Elilily
④TY027(シンガポール)
Tychan
⑤CT-P59(シンガポール)
Celltrion
⑥BRII-196(中国/アメリカ)
Brii Bio/TSB Therapeutics/Tsinghua University(清華大学)/The 3rd People’s Hospital of Shenzhen(深圳市第三人民医院)
⑦BRII-198(中国/アメリカ)
Brii Bio/TSB Therapeutics/Tsinghua University(清華大学)/The 3rd People’s Hospital of Shenzhen(深圳市第三人民医院)
⑧SCTA01(中国)
Sinocelltech Ltd(神州细胞)/Chinese Academy of Sciences(中国科学院)
参考
「Clinical studies evaluating anti-SARS-CoV-2 monoclonal antibodies」
これらのmAbの投与が、ADEの原因につながる不完全抗体を生みうるかどうか?については、今後の研究が必要のようである。
ちなみに、デング熱については、哺乳類細胞で産生されたmAb(mammalian-cell-produced)は、ADE誘発の懸念があるが、植物細胞で産生されたmAb(plant-based)は、ADE誘発の危険性は少ない、との研究結果はすでにある。
(3)ADE誘発は細胞性免疫の誘導失敗が原因
では、どうやってADE誘発リスクをコントロールするのか?
免疫は、そのメカニズムによって2つに大別される。
1つは、液性免疫と呼ばれ、ワクチンの接種や過去の感染によって体内で作られる抗体が活躍する。
抗体には、病原体などに結合することで感染力や毒性を失わせる作用を持つものや、マクロファージなどの免疫細胞による取り込み・処理を助けるものなどがある。
2つは、細胞性免疫と呼ばれ、マクロファージやキラーT細胞といった免疫細胞が病原体や病原体に感染した細胞を直接取り込み(貪食と呼ばれる)、処理することで体を守るシステムである。
細胞性免疫はマクロファージ、キラーT細胞等免疫細胞が感染細胞を貪食で守る。
ADE誘発は、新しいウイルスが、液性免疫の機序を利用して、過去に出来ていた「中和能力はないが吸着能力のある抗体」と複合体を形成し、細胞の中に入り込んだ。
過去の液性免疫の存在が、ADE現象を生み出すもともとの原因であるともいえる。
その次に、免疫細胞であるマクロファージの可逆的極性化特性を利用して、マクロファージのシグナル特性を、ダウンレギュレートの方向に替え、ウイルスの複製環境を整えた。
この後者のマクロファージの特性を変えさせたのは、細胞性免疫の誘導に失敗したためであり、そこにADE現象は起こっている。
ヘルパーT細胞のうち
細胞性免疫の調節がTh1
抗体産生など液性免疫の調節がTh2
で行われるが、ADE現象は、Th1のアップレギュレーションによって、腫瘍壊死因子であるTNF-αの増強を許してしまっていることによる。
ADE現象の発生を回避するには、
細胞免疫の調整を司るTh1と
液性免疫の調整を司るTH2と
のバランスが重要になってくる。
これらの宿主にとっての好ましくない免疫応答が「外生(液性免疫)と内生(細胞性免疫)との二段階」で増強される現象が生じてくる。
図 内生(細胞性免疫)ADE
機序
①抗原と抗体との複合体は、免疫細胞上のFc領域へ結合後、細胞内に内在化
②Th1サイトカインのIL2、TNF-α、IFN-γ を減少させる。
Th2サイトカインのIL-10、IL-6、PGE-2、INF-α を増加させる。
結果、STAT経路を阻害し、免疫応答レベルを低下させる。
③抗ウイルス応答の抑制により、結果として、ウイルス複製は増加。
図 外生(液性免疫)ADE
機序
①抗原&抗体の複合体形成
②複合体が免疫複合体と結合し、補体経路を活性化
③補体フラグメントC4bとC2aとが動員され、C3転換酵素を生成、次にC3が加水分解され、C3bが生成
④C3bは、ウイルスに結合し、CR保有細胞の受容体(CR)を補う
⑤細胞の溶解をし、疾患の悪化を促進させる。
上記図は「Viral-Induced Enhanced Disease Illness」より引用
3.ADEを誘発させない新型コロナウイルスワクチンの条件とは?-終わりに-
ここで、ワクチン接種によって十分な中和抗体が出来るか?が問題となる。
ADE応答は抗体価に依存する。
抗体が低濃度で存在の場合はADEが発生するが、高濃度で存在の場合にADEが発生しない。
また、中和抗体の親和性(結合力)が低い場合にも、ADE現象は発生する。
理 想 的 なワクチンは、SARS-CoV-2 に対して特異的かつ効率的に作用する中和抗体を産生するものでなければならない。
しかし、一定のワクチンの中には、中和抗体の力価が不十分であるために、抗体依存性増強(ADE)が引き起こされる可能性があるワクチンもありうる。
また、ADEと似た症状に「ワクチン過敏反応」(VAH)があるが、これとADE現象と混同しない対応が求められる。
ワクチン抗体産生が不十分で産生された抗体は、新しい感染ウイルスとが結合し、免疫複合体を生成し、Fc受容体経由で、細胞の中に進入する。
十分な力価のないワクチンで産生された抗体、または再感染ウイルスと免疫複合体を作り、ACE2受容体経由でなく、FcγR(Fcガンマ受容体)経由で、マクロファージによる貪食作用で細胞内に進入する。
これらのことから言えば、理想的な新型コロナウイルスワクチンは、「抗体が関与する液性免疫を誘導することなく、細胞性免疫を優位に誘導するワクチン」ということになる。
ワクチン設計にあたり、中和抗体と強力なT細胞性免疫の両方を誘導でき、かつ、ADE誘導性、非中和性、または弱中和性のエピトープを除外することが必要になりうる。
また、中和抗体の出来にくい低力価のワクチンはADE誘発を招きかねず、そのためには、最初のワクチン接種で高力価を保証するための、アジュバントの開発も必要となりうる。
ある識者は、
「S蛋白質に対する不完全な免疫(抗体)が誘導されれば、ADEが起こる可能性がある。
細胞性免疫を誘導するワクチンの開発は、(ADEを防ぐための)1つの手段になり得る。
ウイルスのS蛋白質のうち、感染において特に重要な役割を担う一部の領域をターゲットにしたワクチンを開発することで、不要なエピトープの認識することなく、そのことで、不要な不完全S抗体を生むこともなくなる」
と指摘している。
以上
参考
現在開発中の新型コロナウイルスワクチン一覧
現在開発中のモノクローナル抗体一覧
参考文献
「Antibody-Dependent Enhancement (ADE) Phenomenon Could Be Caused By SARS-COV-2 Antibody」(2020/09/29)
「Immunoglobulin G2 Antibody as a Potential Target for COVID-19 Vaccine」(2020/09/29)
「Immune Response to COVID-19: Can We Benefit from the SARS-CoV and MERS-CoV Pandemic Experience?」 (2020/09/18)
「Another study by researchers from Nanjing University Medical School 」 (2020/09/11)
「Antibody-dependent enhancement and SARS-CoV-2 vaccines and therapies」 (2020/09/09)
「Antibody-dependent enhancement of Coronavirus」(2020/09/07 )
「Immunological considerations for COVID-19 vaccine strategies」 (2020/09/04)
「Four scenarios on how we might develop immunity to Covid-19」 (2020/08/26)
「Structural and dynamical analysis of integrated human/SARS-CoV-2 metabolic models present novel treatment strategies against COVID-19」 (2020/08/14)
「Deciphering SARS-CoV-2 Virologic and Immunologic Features」 (2020/08/14)
「COVID 19 Vaccines: Should we fear ADE? 」 (2020/08/12)
「The role of IgG Fc receptors in antibody-dependent enhancement」(2020/08/11)
「Presence of Genetic Variants Among Young Men With Severe COVID-19」 (2020/07/24)
「Immunity 101 for COVID-19」 (2020/07/18)
「A perspective on potential antibody-dependent enhancement of SARS-CoV-2」 (2020/07/13)
「COVID-19 Coronavirus Vaccine Design Using Reverse Vaccinology and Machine Learning」 (2020/07)
「Implications of antibody-dependent enhancement of infection for SARS-CoV-2 countermeasures」 (2020/06/05)
「Impact of immune enhancement on Covid-19 polyclonal hyperimmune globulin therapy and vaccine development」 (2020/05/01)
「What immunity to Covid-19 might actually mean」(2020/04/23)
「The potential danger of suboptimal antibody responses in COVID-19」(2020/04/21)
「Coronavirus could attack immune system like HIV by targeting protective cells, warn scientists」(2020/04/18)
「SARS-CoV-2 and COVID-19 in older adults: what we may expect regarding pathogenesis, immune responses, and outcomes」 (2020/04/10)
「RETRACTED ARTICLE: SARS-CoV-2 infects T lymphocytes through its spike protein-mediated membrane fusion」 (2020/04/07)
「COVID-19: Immunology and treatment options」 (2020/04)
「Hypothesis for potential pathogenesis of SARS-CoV-2 infection–a review of immune changes in patients with viral pneumonia」 (2020/03/18)
「Immune responses in COVID-19 and potential vaccines: Lessons learned from SARS and MERS epidemic 」 (2020/03)
「Coronavirus infections and immune responses」 (2020/01/25)
「Viral-Induced Enhanced Disease Illness」(2018/12/05)
「Molecular Mechanism for Antibody-Dependent Enhancement of Coronavirus Entry」(2019/12/04)
「FcγRs Muscle Up Dendritic Cell Cross-Presentation」 (2013/01/09)
「Viral Innate Immune Evasion and the Pathogenesis of Emerging RNA Virus Infections」 (2019/10/18)
「Molecular Mechanism for Antibody-Dependent Enhancement of Coronavirus Entry」 (2015)
「Antibody Dependent Enhancement」 (2014)
「Antibody identification」 (?)
「SARS-CoV-2 (COVID-19) S1 protein (D614G), His Tag」 (?)
「Immunology Research Scope」 (?)
「免疫グロブリンFc受 容体を介 した免疫抑制機構」(2000)
「フローサイトメトリー (FCM) / FACS 抗体は、Fc受容体ブロッキングによって検証されます」(?)
以上
以下、メモ的に、順を追って詳述してみたい。
1.ADE(抗体依存性感染増強)現象とは何か?
(1)ADE(抗体依存性感染増強)とは?
ADE(抗体依存性感染増強)(antibody-dependent enhancement)とは
最初に獲得した不完全中和抗体を介し、その中和力はないがFc受容体への吸着力はあるという特性を利用(抗体依存性)して、
次に侵入した抗原(ウイルス)が、不完全中和抗体と複合体を組み、Fc受容体経路で細胞に入り込み、
細胞内で複製力を高める(感染増強)現象
をいう。
具体的には、
①新しい抗原(ウイルス)が、過去に出来ている「中和能力はないが吸着力のある不完全な中和抗体」と結びつき、
②新しい抗原と既成の不完全中和抗体との複合体が出来、
③単球またはマクロファージなどの免疫細胞を呼び寄せる不完全抗体の力を利用しながら、マクロファージの飲作用を利用して、Fc受容体を通過して細胞の中にはいりこみ、
④いったん細胞の中に入り込んだら、ウイルスの複製がうまくいくように、マクロファージのシグナル特性を、上方制御と下方制御の組み合わせで変えさせながら
ウイルス複製環境を増強していく
現象である。
このように、ADE(抗体依存性感染増強)現象は、本来は、宿主を保護するために作られたはずの抗体が、不完全な中和能力のものであったがために、再感染時において、今度は、宿主を助けるのではなく、ウイルスの複製を助ける側に回ってしまうという、きわめて、宿主にとってリスクの多い現象である。
ましてや、そのリスクの多い再感染が、本来は、宿主を救うはずのワクチン接種によっても、もたらされる危険性がある、ということである。
(2)抗体依存性感染増強(ADE)現象の歴史
ADE現象が認識された最初は、1964年のホークス(Hawkes RA. )による報告であった。
(参考 Hawkes RA. 「Enhancement of the infectivity of arboviruses by specific antisera produced in domestic fowls. 」Aust. J. Exp. Biol. Med. Sci.42, 465–482 (1964))
ホークスはこの報告書の中で、アルボウイルス(arbovirus)の感染力が、特定の抗血清の作用によって、見かけの感染力より増加していると指摘した。
アルボウイルスは、節足動物媒介性ウイルスともいわれ、蚊やダニなどの節足動物を介して,吸血 により脊椎動物に伝播されるウイルスである。
フラビウイ ルス科,トガウイルス科,ブニヤウイルス科,レオウイル ス科の多岐に属するウイルスである。
日本脳炎ウイルス(Japanese encephalitis virus)、西ナイルウイルス(West Nile virus)、ゲタウイルス( Getah virus)、マレーバレー脳炎ウイルス(Encephalitis Virus, Murray Valley)、などがあげられている。
その後、1960年から1970年にかけて、同様の現象は、フラビウイ ルス科ウイルス感染を中心にして報告されてきた。
そのほか、
アルファコロナウイルス属では「猫伝染性腹膜炎ウイルス(FIPV)」
に、
ベータコロナウイルス属では「SARS-CoV-1」と「MERS」
に
同様のADE現象が見られている。
今回発生している新型コロナウイルスは、SARSコロナウイルスやMERSコロナウイルス と同じ「コロナウイルス科ベータコロナウイルス属」に分類されている。
参考「動物とヒトのコロナウイルスー2019新型コロナウイルスの流行を受けてー」
また、新型コロナウイルスは「エンベロープを持つRNAウイルス」でもある。
「エンベロープを持つRNAウイルス」には、新型コロナウイルスが含まれるコロナウイルス科のほかに、フラビウイルス科などが含まれる。
参考「エンベローブ(ウイルス)」
フラビウイルス属は、73のウイルスがあり、そのうち、ヒトの重篤疾患に関与しているウイルスが約40ある。
フラビウイルス属は、共通の核酸ゲノム構造(正極性、約11kilobaseの一本鎖RNA)、組織(3つの構造たんぱく質、7つの非構造タンパク質)、粒子サイズ(約500オングストローム;Å)を有している。
この中で、ADE現象に関与しているフラビウイルス属ウイルスとしては、
先に記述した、1964年にホークスが報告している「アルボウイルス(arbovirus)」
や
西ナイルウイルス(West Nile virus)、
デング熱ウイルス(Dengue virus)、
ダニ媒介性脳炎ウイルス(Tick-borne encephalitis virus)、
黄熱病ウイルス(Yellow fever virus)、
ジカウイルス( Zika virus)
などがある。
そのほか
ヒト免疫不全ウイルス(HIV)(Human Immunodeficiency Virus)、
RSウイルス(RSV)(respiratory syncytial viru)、
エボラウイルス(Ebolavirus)、
インフルエンザウイルス(influenzavirus)、
口蹄疫ウイルス(FMD)(Foot-and-mouth disease virus)、
ネコ伝染性腹膜炎ウイルス(FIPV)(feline infectious peritonitis)、
ブタ繁殖・呼吸器障害症候群ウイルス(PRRSV)(Porcine reproductive and respiratory syndrome)
にも、ADE現象の発生がこれまで報告されている。
それ故に、新型コロナウイルスにも、ADE現象が懸念されているのである。
なお、新型コロナウイルスの祖先ウイルスとみられているヒトコロナウイルスにはこれまで、下記のものが発見されている。
αコロナウイルス(γ祖先ウイルスに遠い)-HCoV-229E (1960年代に発見)、HCoV-NL63(2000年代に発見)、
βコロナウイルス(γ祖先ウイルスに近い)-SARS-CoV-2(2019年11月発見)、MERS-CoV(2012年9月発見)、SARS-CoV(2002年11月発見)、HCoV-OC43 (1960年代に発見)、HCoV-HKU1(2000年代に発見)
ADEはウイルスが抗体を利用し細胞侵入をはかり、Fc受容体との相互作用を介し細胞応答を変化させる現象であるが、このことについては、すでに2014年にスコット・B・ハルステッドが他ウイルスにつき下記論文を書いている。
「Pathogenic Exploitation of Fc Activity」
デング熱については、初回の感染時と異なる血清型のウイルスに再感染した場合に、ADE現象が発生することが報告されており、一方、ネココロナウイルスについては、初回の感染時と同じ血清型のウイルスに再感染した場合に、ADE現象が発生することが報告されている。
また、原因未解明のリーシュマニア症(leishmaniasis)(サシチョウバエの吸血により発症)にも、ADE現象が発生するといわれている。
(3)ADE(抗体依存性増強)の機序はどうなっているのか?
A.コロナウイルスが細胞内に入り込む二つの経路
コロナウイルスなど、エンベローブを持つウイルスの細胞侵入過程には次の二経路がある。
①ACE2受容体経路(エンドソーム/リソソーム経路)(endosomal pathway)
②Fc受容体経路(エンドサイトーシス経路)
このうち、ADE現象がみられるのは、②の「Fc受容体経路」である。
図1.ACE2受容体経路
図2.Fc受容体経路
上記2図は「COVID-19: Immunology and treatment options」(SusannaFelsensteinaJenny A.HerbertbPaul S.McNamarabChristian M.Hedrich)を参照
B. ADE誘発は IgG→FcγR(Fcガンマ受容体)経路で起きる
FcγR(Fcガンマ受容体)経由で細胞内侵入のウイルスへの免疫応答機序は下記のとおりである。
①過去に出来た不完全な抗体の存在
宿主内には、過去の感染や接種で産生された不完全な抗体(中和能力はないが、細胞への吸着能力がある抗体)が、既にある。
②新しい抗原と過去の不完全抗体との結合
新しく細胞内に進入しようとしている抗原(新しいウイルス粒子)は、この不完全抗体と結びつき、抗原と抗体との複合体を作る。
③免疫細胞のマクロファージのFc受容体が抗原抗体複合体を認識&結合
免疫細胞であるマクロファージの表面には、Fc受容体があり、不完全中和抗体(免疫グロブリンIgG)の複合体のFc領域を捕らえる分子が出ている。
抗原・抗体の複合体から、抗体の一部を受容体が認識し、結合する。
こうして、過去の不完全な中和抗体が橋渡しをする形で、抗原(新しいウイルス)&過去に産生の不完全中和抗体(免疫グロブリンIgG)の複合体が、マクロファージの表面上のFcγ受容体(Fcガンマ受容体)に吸着&結合する。
抗原(ウイルス)は、過去に出来た不完全抗体と結合したまま、マクロファージの中に取り込まれ、このことで、マクロファージ自体がウイルスに感染してしまう。
低親和性(結合力が弱い)のFcγ受容体(Fcガンマ受容体)は、オプソニン化(抗原に抗体や補体が結合することにより抗原が食細胞に取り込まれやすくなる現象)が促進され、活性化する。
Nikolai Eroshenko等によれば、最初の感染によって誘発された抗体を、新たに侵入の抗体をIgG抗体&Fcガンマ受容体へ、免疫細胞へのウイルスの侵入を可能にするためブリッジするので、ウイルスの向性が変化するためとしている。
既存の抗体には、感染中和の力はないが、Fcガンマ受容体を介しての抗原(ウイルス)の取り込みを誘発する力がある。
そして、既存の抗体は、マクロファージなどの免疫細胞のターゲットとしての機能を果たす。
ウイルスは、宿主の持つ過去に出来た不完全な抗体を「トロイの木馬」として、免疫細胞のターゲットとして差し出し、免疫細胞の食作用のプロセスをだまして利用し、細胞内に入り込む。
④マクロファージがエンドサイトーシス(貧食作用)により複合体を飲み込む
マクロファージはエンドサイトーシス(貧食作用)によって、これら複合体を細胞内に飲み込む。
新しいウイルスは細胞内に入り込み、細胞内のウイルス・ビリオンを増やす(外生的ADE)(B細胞・液性免疫の段階での増強)
⑤本来のマクロファージの機能は?
本来は、マクロファージは次の三つの機能を持つ。
食作用(抗原-ウイルス-を摂取)
↓
抗原提示(食作用によって取り入れた異物を分解して細胞表面に表出させる。これが抗原提示。これがシグナルとなって、ヘルパーT細胞を活性化。)
↓
活性化(マクロファージはT細胞の生産するサイトカインを受け取ることにより活性化)
⑥そして、補体がマクロファージを補助しウイルスを加水分解するはずだが?
この免疫細胞(マクロファージ)の貧食作用を補助するものとして補体があり、マクロファージが食作用で取り込んだウイルスを、補体が、加水分解していく。
本来は、抗原と複合体を形成したIgGにタンパク質複合体C1qが結合し、それをトリガーに「C1→C4→C2→C3b」の経路(古典的経路)でタンパク質分解酵素の生成で、エンべロープウイルスの周りを囲んでいる脂質層の蛋白質を加水分解していく。
なお、第三成分 (C3)が分解されるまでの補体系の活性化には、次の三つのプロセスがある。
古典経路=抗体が関与。補体第一成分 (C1) がC1qを介して結合。
副経路(第二経路)=抗体は関与しない。幾つかの成分の関与でC3が活性化。
マンノース結合レクチン(MBL)経路=MBL(またはMBP (マンナン結合蛋白))が病原体表面の糖鎖に結合して活性化。
⑦補体のエフェクター機能発動までの機序
ADE現象においては、具体的には、以下の図で
a.b.抗原・抗体の複合体にC1qが結合→c.補体のC3.C3a.C5aが活性化、膜侵襲複合体(MAC)が活性化→d.e. IgG Fc受容体は、骨髄細胞上のFcγRに結合し、エフェクター機能を発動させる。→
C3とC5aは骨髄細胞による食作用を促進→d.マクロファージ&樹状細胞による抗体依存性食作用が起きる。→e.ナチュラルキラー(NK)細胞により抗体依存性細胞傷害が媒介。→f.食細胞によるウイルス&ウイルス感染細胞の取り込み後、抗原提示、抗ウイルスT細胞活性化
参考「A perspective on potential antibody-dependent enhancement of SARS-CoV-2」
⑦マクロファージがウイルスに感染し、本来発すべきシグナルを変質させる
ところが、ウイルスに感染したマクロファージは、本来のシグナルを変質させていく。
本来は、複合体を認識したマクロファージは、抗原提示のシグナルを出し、飲み込まれたウイルスは、補体の力を借りて、免疫細胞内で破壊&排除されるはずであるが、細胞内に入ったウイルスがマクロファージに感染(免疫細胞の感染)し、そのマクロファージの出すシグナルを変質させ、自然免疫を抑制させていく。
このマクロファージのシグナルの可逆的な変質を「マクロファージの極性化(Polarization)」という。
マクロファージのシグナル機能が可逆的な極性化によって再プログラム化され、それまでの炎症性細胞から、調節性細胞へ移行していく。
(内生的ADE)(T細胞・細胞性免疫の段階での増強)
⑧マクロファージのシグナル機能の極性化による自然免疫の抑制
マクロファージのシグナル機能の極性化によって、自然免疫は抑制され、結果、ウイルス産生増加は促進される。
インターロイキン10(IL-10)産生増加
ヘルパーT細胞のTh2細胞応答を偏らせる
⑨マクロファージの抗原提示シグナルの変質
マクロファージの出す抗原提示のシグナルにより
T細胞の中のヘルパーT細胞(リンパ球)に伝達。
T細胞活性化で、インターロイキンなどのサイトカインを生産・放出するが、マクロファージのシグナルの変質で、通常とは異なったものとなる。
すなわち
抗原提示のシグナルを送り、
T細胞のなかのヘルパーT細胞(リンパ球)に伝達、
T細胞活性化で、インターロイキンやリンフォカイン等のサイトカインを生産・放出させる。
⑩その結果は?-シグナルの変質化による免疫応答の遅延化で、ウイルス複製環境は増強へ-
シグナルのアップレギュレーションとダウンレギュレーションの相互作用で、免疫応答が遅延してくる。
ここで、もう一度、図2の「Fc受容体経路」に基づいてみてみよう。
マクロファージで取り込まれた抗原&抗体の複合体は、マクロファージの伝達シグナル特性を変えさせ、
①マクロファージのⅠ型IFNシグナル伝達(図のcとe)を阻害(ダウンレギュレーション)
②炎症性サイトカインIL-1.IL-6を発現可能(アップレギュレーション)
③TNFαの発現(図のf)増強(アップレギュレーション)
となり、
結果
Th1サイトカインIL2、TNF-α、IFN-γ減少
Th2サイトカインIL-10、IL-6、PGE-2、INF-α増加
となり、STAT転写因子経路を阻害し、
総合としては、ウイルス・ビリオンの複製環境体制を増強
C.抗原と不完全中和抗体との複合体が出来るまで
a.Fc受容体とは?
抗体のFc領域が担う機能をエフェクター機能という。
Fc領域はそれぞれに特異的なFc受容体に結合し,Fc受容体をもつ細胞を活性化したり,抗体の細胞間トランスポートに働く.
Fc受容体は、
マクロファージや単球などの食細胞、
好中球や好酸球などの顆粒球、
自然免疫系(ナチュラルキラー細胞)や適応免疫系(B細胞など)のリンパ球
など、
免疫系の多くの細胞に見られる。
Fc受容体は、抗体に結合することを可能にする。
抗体に結合し、食細胞の活性化によって、細胞が抗原を排除するのを助ける。
Fc受容体は、Bリンパ球、マクロファージ、ナチュラルキラー細胞、好中球、マスト細胞などの細胞の表面上に存在する。
b. Fc受容体に対応する抗体
Fc受容体に対応する抗体の構造は
Y字型をしており、
F(ab)「アーム」という2本の部位は、抗原への結合領域
Fc「テイル」という1本の部位は
①Fc受容体への結合領域
と
②Fc領域
からなる。
c.5つの免疫グロブリン分子に対する、それぞれ5つのFc受容体
Fc「テイル」の受容体となるFc受容体には次の5つのサブクラスがあり、
5つの免疫グロブリン分子に対しそれぞれ5つのFc受容体
が対応している。
免疫グロブリン分子→Fc受容体
①IgG→FcγR(Fcガンマ受容体)
(IgGは血中で最も豊富に認められる抗体。産生されるサイトカインの種類によって、4種類のIgGのサブクラスがある。
IgG1およびIgG3は、食細胞のFc受容体に高い親和性を示す。
食細胞のFc受容体経由オプソニン化(抗原に抗体や補体が結合することにより抗原が食細胞に取り込まれやすくなる現象)に有効。
Fc領域を介してナチュラルキラー細胞にも結合することができる。
抗体依存性の細胞毒性に寄与)
②IgE→FcεR(Fcイプシロン受容体)
(IgEは粘膜表面、血液、組織において機能)
③IgD→FcδR(Fcデルタ受容体)
(IgDはほとんどのBリンパ球の表面に存在)
④IgM→FcμR(Fcミュー受容体)
(IgMは新生児が最初に合成する免疫グロブリン)
⑤IgA→FcαR(Fcアルファ受容体)
(IgAは中和抗体としての作用)
参考「抗体の仕組みと種類を理解しよう」
d.ADE誘発に関わるFcγR(Fcガンマ受容体)に対する免疫グロブリン分子はIgG
ADE誘発に関係のFc受容体はFcγR(Fcガンマ受容体)で免疫グロブリン分子IgGの受容体である。
なお、新生児の抗体であるIgMも、ADEを誘発しうるとの説がある。
結合リガンド(特異結合物質)
は、上から
→抗原(Antigen)と結合
→糖鎖修飾誘導体(Glycan receptor)と結合
→Fc受容体(FcR)と結合
参考「Pharmacokinetics of monoclonal antibodies and Fc-fusion proteins」
免疫応答を引き出す新しいウイルス抗原は既存の抗体の免疫グロブリン分子IgGと結合し、抗原と抗体の複合体完成でIgGのY字の根元部分で受容体に結合する。
ADE現象を誘発する抗体は
F(ab)の結合力が弱いが、Fcの結合力が強いので、免疫細胞を捕らえてしまう。
これには、Fc部位の糖化状態が、関係しているとの説もある。
IgG応答は、ウイルス検出後、7日後から観察され、長期間にわたり、高いレベルで続くことがある。
なお、上記「糖鎖修飾誘導体(Glycan receptor)と結合」がADE現象に与える影響については、研究文献が少ないが、例えば、代表的な翻訳語修飾としてある「リン酸化」や「フコース」がウイルス核内移行やサイトカイン産生などにどう影響しているのか?については、興味のあるところである。
D. ACE2受容体経路は、ADEには無関係なのか?
ACE10経路の受容体は、スパイクタンパク質主体の受容体ではあるが、単球・マクロファージの受容体としては、発現が低い。
しかし、スパイクタンパク質の表面のエピトープ(抗体が認識する抗原の一部分)の異なる部分の認識によって不完全中和抗体が出来ている可能性は十分ある。
ましてや、現在世界で開発の新型コロナウイルスワクチンは、ほとんどが、ウイルスのスパイクタンパク質を標的にしたものである。
十分な力価を持ち、十分な中和抗体を形成するワクチンなら、ACE2受容体へのS蛋白質結合をブロックしうる。
本来は、スパイク蛋白質とその免疫は防御免疫応答を誘発しうるはずである。
しかし、中途半端な力価のワクチンが交差反応を起こして、不十分な中和抗体を形成し、これが、新規のウイルスの抗原&抗体複合体形成のための標的になり、FcγR(Fcガンマ受容体)経由で細胞に侵入してくる機会は十分想定しうる。
これらの複合体は、抗体力はないが、受容体への吸着力はある。
抗スパイクIgGの存在が、マクロファージ/単球におけるFcγRIIシグナル伝達を介して、免疫応答を歪めることで、ADE感染増強に関連している可能性は考えられる。
E.過炎症の指標
ADE現象の結果として起こりうるサイトカインストームによる過炎症(またはさらに超炎症;Hyper inflammation)の指標として、次のものがあげられている。
白血球(WBC)数
好中球
リンパ球数
好中球/リンパ球比(NLR)
血小板/リンパ球比(PLR)
このうち、好中球/リンパ球比(NLR)は、全身性炎症のマーカーとして、注目されている。
この比率が3を超えると、リンパ球の相対的減少状態にあるとされ、細胞性免疫の低下とみなされる・
新型コロナウイルス感染による重症化の指標とされている。
2.「抗体依存性感染増強」(ADE)現象を防ぐには?
(1)ADE誘発の最大の原因は、過去から存在していた不十分な中和抗体の存在
これまで見たように、ADE誘発の最大の原因は、次のような不完全中和抗体の存在である。
①過去の中途半端なウイルス感染や質の悪いワクチン接種によって出来た力価の不十分な中和抗体
②異種ウイルスに対する交差反応によって出来た不十分な中和抗体
③スパイクタンパク質の表面のエピトープの異なる部分の認識によって、過去に出来た不完全中和抗体
④他のコロナウイルス(先祖ウイルスかもしれないコウモリコロナウイルス)のプライミング(初回感染曝露時に免疫系を賦活するための予備刺激or準備刺激or初回刺激)によって発生した不完全中和抗体
⑤T細胞のTh2応答を刺激したかもしれない、過去に接種の麻疹ワクチンによって産生されたワクチン抗体
⑥血清型が異なるために、単に亜中和されただけの不完全中和抗体
⑦風土病化した不顕性感染のウイルスに曝露されてきた宿主が持っている不完全中和抗体
の存在である。
これらの存在が、ウイルスへの再感染やワクチンの再接種によって、ADE現象を生み出す要因になっている。。
これらは、中和抗体を作る能力はないが、Fc受容体に結合する能力がある。
そのために、ADE現象を引き起こしてしまっている。
(2).モノクローナル抗体とADEの関係は?
現在、新型コロナウイルスに対するモノクローナル抗体(mAb)としては、次の8つが研究されており、それぞれ臨床試験の段階にある。
①REGN10933+REGN10987(アメリカ)
Regeneron/N IAD
②LY3819253(LY-CoV555)(カナダ/アメリカ)
AbCellera/Eli Lilly
③JS016(中国/アメリカ)
Junshi Biosciences (上海君实生物医药科技股份有限公司)/Institute of Microbioligy.Chinese Academy of Sciences/Elilily
④TY027(シンガポール)
Tychan
⑤CT-P59(シンガポール)
Celltrion
⑥BRII-196(中国/アメリカ)
Brii Bio/TSB Therapeutics/Tsinghua University(清華大学)/The 3rd People’s Hospital of Shenzhen(深圳市第三人民医院)
⑦BRII-198(中国/アメリカ)
Brii Bio/TSB Therapeutics/Tsinghua University(清華大学)/The 3rd People’s Hospital of Shenzhen(深圳市第三人民医院)
⑧SCTA01(中国)
Sinocelltech Ltd(神州细胞)/Chinese Academy of Sciences(中国科学院)
参考
「Clinical studies evaluating anti-SARS-CoV-2 monoclonal antibodies」
これらのmAbの投与が、ADEの原因につながる不完全抗体を生みうるかどうか?については、今後の研究が必要のようである。
ちなみに、デング熱については、哺乳類細胞で産生されたmAb(mammalian-cell-produced)は、ADE誘発の懸念があるが、植物細胞で産生されたmAb(plant-based)は、ADE誘発の危険性は少ない、との研究結果はすでにある。
(3)ADE誘発は細胞性免疫の誘導失敗が原因
では、どうやってADE誘発リスクをコントロールするのか?
免疫は、そのメカニズムによって2つに大別される。
1つは、液性免疫と呼ばれ、ワクチンの接種や過去の感染によって体内で作られる抗体が活躍する。
抗体には、病原体などに結合することで感染力や毒性を失わせる作用を持つものや、マクロファージなどの免疫細胞による取り込み・処理を助けるものなどがある。
2つは、細胞性免疫と呼ばれ、マクロファージやキラーT細胞といった免疫細胞が病原体や病原体に感染した細胞を直接取り込み(貪食と呼ばれる)、処理することで体を守るシステムである。
細胞性免疫はマクロファージ、キラーT細胞等免疫細胞が感染細胞を貪食で守る。
ADE誘発は、新しいウイルスが、液性免疫の機序を利用して、過去に出来ていた「中和能力はないが吸着能力のある抗体」と複合体を形成し、細胞の中に入り込んだ。
過去の液性免疫の存在が、ADE現象を生み出すもともとの原因であるともいえる。
その次に、免疫細胞であるマクロファージの可逆的極性化特性を利用して、マクロファージのシグナル特性を、ダウンレギュレートの方向に替え、ウイルスの複製環境を整えた。
この後者のマクロファージの特性を変えさせたのは、細胞性免疫の誘導に失敗したためであり、そこにADE現象は起こっている。
ヘルパーT細胞のうち
細胞性免疫の調節がTh1
抗体産生など液性免疫の調節がTh2
で行われるが、ADE現象は、Th1のアップレギュレーションによって、腫瘍壊死因子であるTNF-αの増強を許してしまっていることによる。
ADE現象の発生を回避するには、
細胞免疫の調整を司るTh1と
液性免疫の調整を司るTH2と
のバランスが重要になってくる。
これらの宿主にとっての好ましくない免疫応答が「外生(液性免疫)と内生(細胞性免疫)との二段階」で増強される現象が生じてくる。
図 内生(細胞性免疫)ADE
機序
①抗原と抗体との複合体は、免疫細胞上のFc領域へ結合後、細胞内に内在化
②Th1サイトカインのIL2、TNF-α、IFN-γ を減少させる。
Th2サイトカインのIL-10、IL-6、PGE-2、INF-α を増加させる。
結果、STAT経路を阻害し、免疫応答レベルを低下させる。
③抗ウイルス応答の抑制により、結果として、ウイルス複製は増加。
図 外生(液性免疫)ADE
機序
①抗原&抗体の複合体形成
②複合体が免疫複合体と結合し、補体経路を活性化
③補体フラグメントC4bとC2aとが動員され、C3転換酵素を生成、次にC3が加水分解され、C3bが生成
④C3bは、ウイルスに結合し、CR保有細胞の受容体(CR)を補う
⑤細胞の溶解をし、疾患の悪化を促進させる。
上記図は「Viral-Induced Enhanced Disease Illness」より引用
3.ADEを誘発させない新型コロナウイルスワクチンの条件とは?-終わりに-
ここで、ワクチン接種によって十分な中和抗体が出来るか?が問題となる。
ADE応答は抗体価に依存する。
抗体が低濃度で存在の場合はADEが発生するが、高濃度で存在の場合にADEが発生しない。
また、中和抗体の親和性(結合力)が低い場合にも、ADE現象は発生する。
理 想 的 なワクチンは、SARS-CoV-2 に対して特異的かつ効率的に作用する中和抗体を産生するものでなければならない。
しかし、一定のワクチンの中には、中和抗体の力価が不十分であるために、抗体依存性増強(ADE)が引き起こされる可能性があるワクチンもありうる。
また、ADEと似た症状に「ワクチン過敏反応」(VAH)があるが、これとADE現象と混同しない対応が求められる。
ワクチン抗体産生が不十分で産生された抗体は、新しい感染ウイルスとが結合し、免疫複合体を生成し、Fc受容体経由で、細胞の中に進入する。
十分な力価のないワクチンで産生された抗体、または再感染ウイルスと免疫複合体を作り、ACE2受容体経由でなく、FcγR(Fcガンマ受容体)経由で、マクロファージによる貪食作用で細胞内に進入する。
これらのことから言えば、理想的な新型コロナウイルスワクチンは、「抗体が関与する液性免疫を誘導することなく、細胞性免疫を優位に誘導するワクチン」ということになる。
ワクチン設計にあたり、中和抗体と強力なT細胞性免疫の両方を誘導でき、かつ、ADE誘導性、非中和性、または弱中和性のエピトープを除外することが必要になりうる。
また、中和抗体の出来にくい低力価のワクチンはADE誘発を招きかねず、そのためには、最初のワクチン接種で高力価を保証するための、アジュバントの開発も必要となりうる。
ある識者は、
「S蛋白質に対する不完全な免疫(抗体)が誘導されれば、ADEが起こる可能性がある。
細胞性免疫を誘導するワクチンの開発は、(ADEを防ぐための)1つの手段になり得る。
ウイルスのS蛋白質のうち、感染において特に重要な役割を担う一部の領域をターゲットにしたワクチンを開発することで、不要なエピトープの認識することなく、そのことで、不要な不完全S抗体を生むこともなくなる」
と指摘している。
以上
参考
現在開発中の新型コロナウイルスワクチン一覧
現在開発中のモノクローナル抗体一覧
参考文献
「Antibody-Dependent Enhancement (ADE) Phenomenon Could Be Caused By SARS-COV-2 Antibody」(2020/09/29)
「Immunoglobulin G2 Antibody as a Potential Target for COVID-19 Vaccine」(2020/09/29)
「Immune Response to COVID-19: Can We Benefit from the SARS-CoV and MERS-CoV Pandemic Experience?」 (2020/09/18)
「Another study by researchers from Nanjing University Medical School 」 (2020/09/11)
「Antibody-dependent enhancement and SARS-CoV-2 vaccines and therapies」 (2020/09/09)
「Antibody-dependent enhancement of Coronavirus」(2020/09/07 )
「Immunological considerations for COVID-19 vaccine strategies」 (2020/09/04)
「Four scenarios on how we might develop immunity to Covid-19」 (2020/08/26)
「Structural and dynamical analysis of integrated human/SARS-CoV-2 metabolic models present novel treatment strategies against COVID-19」 (2020/08/14)
「Deciphering SARS-CoV-2 Virologic and Immunologic Features」 (2020/08/14)
「COVID 19 Vaccines: Should we fear ADE? 」 (2020/08/12)
「The role of IgG Fc receptors in antibody-dependent enhancement」(2020/08/11)
「Presence of Genetic Variants Among Young Men With Severe COVID-19」 (2020/07/24)
「Immunity 101 for COVID-19」 (2020/07/18)
「A perspective on potential antibody-dependent enhancement of SARS-CoV-2」 (2020/07/13)
「COVID-19 Coronavirus Vaccine Design Using Reverse Vaccinology and Machine Learning」 (2020/07)
「Implications of antibody-dependent enhancement of infection for SARS-CoV-2 countermeasures」 (2020/06/05)
「Impact of immune enhancement on Covid-19 polyclonal hyperimmune globulin therapy and vaccine development」 (2020/05/01)
「What immunity to Covid-19 might actually mean」(2020/04/23)
「The potential danger of suboptimal antibody responses in COVID-19」(2020/04/21)
「Coronavirus could attack immune system like HIV by targeting protective cells, warn scientists」(2020/04/18)
「SARS-CoV-2 and COVID-19 in older adults: what we may expect regarding pathogenesis, immune responses, and outcomes」 (2020/04/10)
「RETRACTED ARTICLE: SARS-CoV-2 infects T lymphocytes through its spike protein-mediated membrane fusion」 (2020/04/07)
「COVID-19: Immunology and treatment options」 (2020/04)
「Hypothesis for potential pathogenesis of SARS-CoV-2 infection–a review of immune changes in patients with viral pneumonia」 (2020/03/18)
「Immune responses in COVID-19 and potential vaccines: Lessons learned from SARS and MERS epidemic 」 (2020/03)
「Coronavirus infections and immune responses」 (2020/01/25)
「Viral-Induced Enhanced Disease Illness」(2018/12/05)
「Molecular Mechanism for Antibody-Dependent Enhancement of Coronavirus Entry」(2019/12/04)
「FcγRs Muscle Up Dendritic Cell Cross-Presentation」 (2013/01/09)
「Viral Innate Immune Evasion and the Pathogenesis of Emerging RNA Virus Infections」 (2019/10/18)
「Molecular Mechanism for Antibody-Dependent Enhancement of Coronavirus Entry」 (2015)
「Antibody Dependent Enhancement」 (2014)
「Antibody identification」 (?)
「SARS-CoV-2 (COVID-19) S1 protein (D614G), His Tag」 (?)
「Immunology Research Scope」 (?)
「免疫グロブリンFc受 容体を介 した免疫抑制機構」(2000)
「フローサイトメトリー (FCM) / FACS 抗体は、Fc受容体ブロッキングによって検証されます」(?)
以上
1.ワクチン開発の状況
A
.
1.AJワクチン(AJ Vaccines)
2.アルティミューン(Altimmune)
準有力(ワクチン)
アデノウイルスベースのNasoVAXワクチン
鼻腔内ワクチン
2月28日にワクチン設計完了
フェーズ1臨床試験は早ければ8月中旬開始
.
3.Applied DNA Science
4.アークトゥルス・セラピューティクス(Arcturus Therapeutics Ltd.)と
デュークNUSメディカルスクール(DukeNUS Medical School)(在シンガポール)
有力(ワクチン)
RNA(STARR)ワクチン
STARRテクノロジーに基づく低廉なワクチンの開発を最短で、が目標。
デュークNUSメディカルスクール開発のプラットフォームを利用。
既に生産段階に入っている。
.
B
.
5.ベイラー医学大学(Baylor College of Medicine)
S1蛋白質ワクチン
RDB蛋白質ワクチン
.
6.Beijing Advaccine Biotechnology( 北京艾棣维欣生物技术有限公司)
Inovio と連携。VGXIも協力。
IN-4800(ワクチン)
第Ⅰ相試験段階
.
7.Beijing Dingcheng Taiyuan Biotechnology(北京鼎成肽源生物技术有限公司)と Betta Pharmaceuticals(贝达药业股份有限公司)
樹状細胞ワクチン(Dendridic Cell Vaccine)
探索研究段階
8.BioNTech
有力(ワクチン)
mRNAベース、BNT162ワクチン。
ファイザー、Fosum Pharmaと提携。
2020年4月下旬に臨床試験に入る。
.
C
.
9.カンシーノ(CanSinoBIO)
有力(ワクチン)
有力(ワクチン)
mRNAワクチン。
探索研究段階
狂犬病のワクチンに成功したドイツの会社。
官民連合の「感染症流行対策イノベーション連合(CEPI)((Coalition for Epidemic Preparedness Innovations)に協力
フェーズⅠ試験は2020年6月か7月
.
16.ドハーティ研究所(Doherty Institute)
20.エビバックス(Epivax)とロスラボ(RossLab)
Ii-Keyペプチドワクチン
ジェネレックス 中国のコンソーシアムが資金提供
2020年6月からヒト実験に入る。
.
25.ゲオバックス(GeoVax)
準有力(ワクチン)
蛋白質ベースのS-Trimerワクチン
グラクソスミスクライン 中国の三叶草生物制药(Clover Biopharmaceuticals)とのコラボレーション
三叶草生物制药のワクチンはRecombinant Subunit ワクチン。
GSKがアジュバントを三叶草生物制药に提供。
アメリカの重点支援企業
.
27.Guanhao Biotech
Z-VacciRNA ワクチン
探索研究段階
.
H
.
28.ヒートバイオロシクス(Heat Biologics)とマイアミ大学(University of Miami)
準有力(ワクチン)
2020年3月3日にプログラム発表
サブユニットワクチン
iBioのFast Pharming Systemを利用して生産。
Agrobacteriumをプラットフォーム
植物(タバコ)由来のワクチン(plant–based vaccine )
2020年2月にプログラム発表
.
30.ImmunoPrecise
ワクチンと中和抗体
PolyTope mAb Therapy
探索研究段階
.
31.インペリアルカレッジ ロンドン(Imperial College London)
INO-4800ワクチン。
北京艾棣维欣生物技术有限公司(Beijing Advaccine Biotechnology)とのコラボレーション
パンデミック・シナリオ用に設計。
第Ⅰ相試験実施
中国→韓国→米国で3,000ドーズ 試験接種
.
34.パスツール研究所(Institut Pasteur)
準有力(ワクチン)
Prezcobixワクチン
アデノウイルスを弱毒化
BARDAと協力
アメリカの重点支援企業
マウス実験済
FDAがファストトラック適用で、パンデミック&全災害準備再認可法(Pandemic and All-Hazards Preparedness Reauthotization Act of 2013(PAHPRA))の「緊急使用可」プロセス適用で、三段階承認プロセスが回避されることを見込んでいる。
2020年9月からフェーズⅠ実験に入る予定。
2021年初頭から緊急用として10億ドーズを供給予定、
.
L
.
37.LineaRx
準有力(ワクチン)
線形DNAワクチン
中和抗体の誘導
ローマ本拠のTakisBiotechと協力
2020年5月か6月にフェーズⅠ実験に
.
M
.
38.Mallinckrodt
INOmax (nitric oxide) 吸入ワクチン
.
39.メディカゴ(Mediacago)
植物(タバコ)ベースワクチン。
前臨床実験中
2020年夏からフェーズⅠ実験へ。
田辺三菱製薬がこれを使ったCOVID-19向けワクチンの非臨床試験を行っている。
2010年8月までにヒトでの臨床試験を開始予定
.
40.Medigen Biotechnology
SARS-COV-2ワクチン
台湾の会社
探索研究段階
.
41.ミガル研究所(MIGAL Galilee Research Institute)
有力(ワクチン)
IBV
経口ワクチン
イスラエルに拠点を置く
.
42.モデルナ(ModernaTherapeutics)
有力(ワクチン)
mRNA-1273ワクチン
コロナウイルスの遺伝子配列確定後42日後に作成。
世界で最も開発が進んでいるワクチン。
動物実験データなしに、2020年3月16日からヒトに接種実験
トライアル研究では健康な成人に3ドーズを28日間間隔で接種。
ワクチン設計は、NIAIDワクチン研究センター(VRC)と共同研究
アメリカの重点支援企業
インペリアルカレッジ ロンドン(Imperial College London)のMucosal Infection and Immunityとも関連。
.
N
.
43. NanoViricides
Anti-SARS
探索研究段階
.
44.ノヴァヴァックス(Novavax)
O
.
45.大阪大学( Osaka University)
準有力(ワクチン)
アンジェス(Anges)
ウイルスを製造に持ち込まないデオキシリボ核酸(DNA)ワクチン
3月26日には非臨床試験開始。
6カ月以内のできる限り早い時期の臨床試験開始を目指す。
.
S
.
46.サノフィ(Sanofi)
準有力(ワクチン)
DNA組み換えワクチン
既に開発のサノフィ SARSワクチンをベースにしたもの、
BARBAと協力
2月18日、BARDAの協力を得て、これまで開発のSARS用ワクチン開発で蓄積の技術をコロナウイルスにも適用するとした。
アメリカの重点支援企業
前臨床実験中
臨床実験開始は2021年3月から8月にかけて
.
47.Shenzhen Kangtai Biological Products
.
48.Sichuan Clover Biopharmceuticals
.
49.Sinopharm Group
.
50.Stermirma Therapeutics & Tongui University
関係の記事or論文サイトへリンク
.
51.Sumagen
.
52.シノバック(Sinovac)
53.Tonix Pharmaceuticals
準有力(ワクチン)
TNX-1800 改変馬poウイルスワクチン
2020年2月に臨床実験のIND申請
.
54.Tonix Pharmaceuticals Holding
有力(ワクチン)
経皮投与の生ワクチンTNX-1800
本来は、アメリカの戦略的国家備蓄用の天然馬痘予防生ワクチン及びサル用予防ワクチンとして開発中だった。
馬のウイルスをベクターとして活用
Southeren Researchと提携
.
55.チューレン大学(Tulane University)
CSLの子会社Seqirusが有するアジュバント技術を大学に提供
Dynavax
2020年3月中旬時点で前臨床段階
.
59.サスカチュワン大学(University of Saskatchewan)
60.バクサート(Vaxart)
準有力(ワクチン)
錠剤投与の経口組み換えワクチン。
探索研究段階
常温保管が出来る。
粘膜粘液を保護。
Emergent Bio Solutions
2020年3月中旬時点で前臨床段階
.
61.ヴァクシル(VAXIL)
準有力(ワクチン)
DNAワクチンと 生弱毒組換え麻疹ウイルス(rMV)ベクターワクチン
インドの会社。
二つのアプローチでワクチン開発。
ウイルスの細胞侵入に関わるタンパク質を標的にしたもの
と
弱毒化した組み換えマシンウイルス(rMV)によるワクチン
コドン最適化蛋白質を発現し、長期中和抗体を誘導
.
.
.
2.抗体・既存薬剤・
阻害剤・治療方法
.
A
.
1.アッヴィ(AbbVie)
有力(阻害剤)
カレトラ(Kaletra)(ロビナビル(Lopinavir)とリトナビル(Ritonavir)との合剤)使用
もともとは、HIV-1プロテアーゼ阻害剤。
1月27日に中国国家健康委員会が、コロナ治療に許可。
.
2.AbCellera Biologics
有力(モノクローナル抗体)
.
3.AIM ImmunoTech
有力(免疫調節薬)
リンタトリモド(Rintatolimod)(商品名アンプリジェン(Ampligen ))使用
上市済
本来は、重度の慢性疲労症候群に向けた免疫調節薬。
ヨハン・ヴォルフガング・ゲーテ大学(Johann Wolfgang Goethe–Universität )と協力
.
4.APEIRON Biologics
有力(組み換え蛋白質)
組換えヒトアンジオテンシン変換酵素(Angiotensin converting enzyme 2 (ACE2) )(rhACE2)使用
本来は、急性肺損傷肺動脈高血圧症のための薬。
.
6.アスクレティスファーマ(Ascletis Pharma)
有力(阻害剤)
ダノプレビル(danoprevir)(商品名 Ganovo)使用
第III相試験(フェーズ III)
本来は、C型肝炎ウイルスプロテアーゼ阻害薬。
併用服用(リトナビル(Ritonavir)+ASC09やタミフル(Oseltamivr))
.
6-2.アストラゼネカ(AstraZeneca)
有力(他用途薬)
アカラブルチニブ(acalabrutinib)(商品名カルクエンス(Calquence))
本来は マントル細胞リンパ腫 の薬。
.
7. Australia Murdoch Children’s Research Institute (Melbourne)
既存のBCG(カルメットゲラン菌(Bacille de Calmette et Guérin))の接種
臨床試験中
.
B
.
8.バイエル(Bayer)
有力(他用途薬)
リン酸クロロキン(Chloroquine Phosphate)(商品名 レゾシン(Resochin))と抗ウイルス薬のレムデシビル(remdesivir)使用。
上市済
リン酸クロロキンは、本来は抗マラリア、抗炎症性を持つキノリン化合物。
北京の病院などで使用。
.
9.北京德丰瑞生物科技有限公司(Beijing Defengrei Biotehnology)
有力(モノクローナル抗体)
InflaRxと提携
BDB-1
第Ⅱ相試験(フェーズ Ⅱ)段階
.
10.舒泰神北京生物製薬(Beijing Staidson Biopharma)とInflaRx
有力(モノクローナル抗体)
IFX-1(抗C5aモノクローナル抗体)使用
有力(阻害剤)
ガリデシビル(Galidesivir)(BCX4430)使用
本来はダニ媒介性脳炎ウイルス阻害剤で、ウイルス複製を妨害するヌクレオシドRNAポリメラーゼ阻害
.
15.BioCubaFarma
有力(抗ウイルス薬)
インターフェロンアルファ2B(商品名 IFNrec)
中国と連携し吉林省Changheber工場で生産。
インターフェロン・アルファ-2Bリコンビナント(IFNrec)という抗ウイルス薬は、キューバと中国の研究チームが共同で開発
.
16.BioXyTran
17.カモスタット(Camostat,INN)
19.セルラリティ(Celularity)とソレント・セラピューティクス(Sorrento Therapeutics)
有力(細胞療法)
同種移植。
ナチュラルキラー細胞(NK細胞)の CYNK-001使用
前臨床試験段階
胎盤由来NK細胞療法
.
20.中外製薬(Chugai Pharmaceutical )と浙江海正薬業股彬有限公司(Zhejiang Hisun Pharmaceutical Co)
ヒト化抗ヒトIL-6受容体モノクローナル抗体のトシリズマブ(Tocilizumab)(ヒト化mAb)使用
本来はインターロイキン-6(IL-6)の作用を抑制し免疫抑制効果を示す分子標的治療薬
.
21.Cocrystal
有力(阻害剤)
プロテアーゼ阻害剤
カンサス州立大学(Kansas State University)と協力
23.CytoDyn
有力(阻害剤)
レロンリマブ(Leronlimab)(PRO 140)(ヒト免疫系のTリンパ球に見られるCCR5受容体を標的とするヒト化モノクローナル抗体)を使用
本来はCCR5阻害薬で、CCR5という第二の受容体とのウイルスの結合を阻害。
.
27.Eli Lilly
有力(抗体)
アブセレラ・バイオロジクス(AbCellera Biologics)との間で、新型コロナウイルス向けの抗体ベースの治験薬を共同開発
.
28.Emergent Bio Solution
有力(抗体)
ポリクローナル抗体 CoVid-HIG.CoVid-EIG
.
29.Enata Pharmaceuticals
抗ウイルス剤
.
30.EUSA Pharma
有力(モノクローナル抗体)
.
F
.
31.富士フイルムホールディングス(FUJIFILM Holdings Corporation)と浙江海正薬業股彬有限公司(Zhejiang Hisun Pharmaceutical Co)
有力(他用途薬)
ファビピラビル( Favipiravir)(商品名 アビガン(Abigan))使用
上市済
本来は抗ウイルス薬で、RNAウイルスのRNA依存性RNAポリメラーゼ(RdRp)を選択的に阻害。
旧 富山化学 開発
中国では、2月17日に治療薬として承認。
.
G
.
I
.
34.ImmunoPreciseAntibodies
有力(モノクローナル抗体)
抗体 AIによる 計算抗体設計
.
35.Impact Bio Medial
有力(抗体)
受容体ACE2 結合化合物
.
36.インペリアルカレッジロンドン(Imperial College London)
粘膜感染と免疫
.
37.InflaRx
有力(モノクローナル抗体)
.
38.Incyteと上海恒瑞医药有限公司(Shanghai Hengrui Pharmaceutical)
有力(モノクローナル抗体)
上海江蘇恒瑞医薬のカムレリズマブ(Camrelizumab)(PD-1を阻害するヒト化モノクローナル抗体)
と
サイモシン(Thymosin)(5Daポリペプチドホルモン( 5-Da polypeptide hormone))
との併用使用
.
39.イノベーション ファーマシューティカルズ(Innovation Pharmaceuticals)
有力(他用途薬)
ブリラシジン(Brilacidin )使用
第Ⅱ相試験(フェーズ Ⅱ)段階
本来は頭頸部がん患者の口内粘膜炎の薬
.
43.ジョンソン&ジョンソン(Johnson&Johnson)
有力(阻害剤)
商品名 プレジコビックス (Prezcobix) (ダルナビル エタノール付加物(Darunavir ethanolate). コビシスタット(
本来はHIVプロテアーゼ阻害剤
ロピナビルやリトナビルと組み合わせ使用
48.Nascent biotech
50.OyaGen
有力(抗ウイルス剤)
広域抗ウイルス剤 OYA1
エボラ熱などのフィロウイルスにも有効
54.Q Biomed と Mannin Research
有力(補助治療)
アンジオポエチン(angiopoietin)タンパク質に注目の血管漏出および内皮機能不全の補助治療(MAN-01)
アンジオポエチン(angiopoietin)は、脈管形成(vasculogenesis)あるいは血管新生(angiogenesis)を促進する糖タンパク質。
両機関は2月4日に提携。
.
R
.
55.Redhill Biopharma
有力(阻害剤)
「Opaganib (ABC294640) 」(商品名 Yeliva )
スフインゴシン キナーゼ2型阻害剤
トリプシン株 セリンプロテアーゼ阻害剤 RHB-1011 RHB-1011
.
.
58.サノフィ( Sanofi S.A.)
有力(他用途薬剤)
59.上海交通大学医学院附属瑞金病院(Shanghai jiaotong university school of medicine Ruijin Hospital)
(他用途阻害剤)
本来はインフルエンザ治療薬として開発された膜融合阻害剤ウミフェノビル( Umifenovir) (商品名 アルビドール(arbidol))を使用
カレトラ(アッヴィ社)、ASO99(アスクレティスファーマ社)、ロピナビル、リトナビル、カリミシン、塩酸ブロムヘキシン、などと併用使用。
瑞金病院では単独使用。
.
60.Sichuan Kelun Pharmaceutical (四川科倫薬業股分有限公司)
S-Protein/ACE2
(ポリペプチド)
前臨床試験段階
プロイン/ACE2 ターゲットのポリペプチド
関係の記事or論文サイトへリンク
.
60-2.塩野義製薬 ほか
低用量プレドニゾロン(アレルギーの治療に使用されるステロイド)とタクロリムス療法(臓器移植を受けた患者に与えられる免疫抑制薬)が新型コロナウイルス感染に有益な影響を与える可能性がある。
.
T
.
61.武田(Takeda)
62.帝人ファーマ(TEIJIN PHARMA LIMITED)
(他用途薬剤)
64.ウィル・バイオテクノロジー(Vir Biotechnology)と薬明生物技術有限公司(WuXi Biologics)
有力(モノクローナル抗体)
ヒトモノクローナル抗体
.
65.VIR Biotechnology

以上